Эволюционные адаптации пчел: сбор нектара, пыльцы и цветочных масел
Цветочные ресурсы: Нектар и его роль в метаболизме. Нектар является основным источником углеводов в рационе пчел, обеспечивая энергию для взрослых особей и служа важным компонентом корма для личинок. Помимо сахаров, этот ресурс содержит аминокислоты, которые могут вносить существенный вклад в азотистый обмен насекомых. Состав нектара варьируется в зависимости от вида растения, адаптируясь под конкретных опылителей. Пчелы собирают его в специальный орган — зобик (crop), где он транспортируется в гнездо.
Процесс поглощения нектара осуществляется с помощью сложного ротового аппарата, называемого пробосцисом. Анатомическое строение этой структуры подробно представлено на рисунках 19-1 — 19-5. Ротовое отверстие располагается на передней поверхности основания пробосциса (рис. 19-1c). Движение жидкости к ротовому отверстию обеспечивается за счет работы максиллярных галей (maxillary galeae), которые формируют защитную оболочку.
У длинноязыких пчел этот механизм дополняется лабиальными щупиками (labial palpi), окружающими глоссу (glossa). Транспортировка нектара происходит благодаря капиллярному эффекту и ритмичным движениям челюстей. Harder (1983) детально описал эти механизмы для рода Andrena, а Snodgrass (1956) представил классическое описание анатомии Apis. Эффективность сбора напрямую зависит от вязкости нектара и морфологии ротовых частей.
Глосса, покрытая множеством волосков, способна модулировать объем удерживаемого нектара за счет их выпрямления и прижатия при втягивании. Форма этих волосков крайне разнообразна: от простых и уплощенных до ланцетных или разветвленных (рис. 59-3, 84-1, 114-2). Эти микроструктуры увеличивают площадь поверхности, способствуя лучшему удержанию жидкости. Эволюция глоссы шла по пути оптимизации сбора ресурсов с цветков различной глубины.
Секреция и сбор цветочных масел. В исключительных случаях растения из семейств Cucurbitaceae, Iridaceae, Krameriaceae, Malpighiaceae, Orchidaceae, Primulaceae, Scrophulariaceae и Solanaceae выделяют цветочные масла вместо нектара. Эти жирные секреты вырабатываются специальными железами — элайофорами. Пчелы-специалисты собирают эти масла, используя специализированные структуры на конечностях. Масла смешиваются с пыльцой и иногда нектаром для создания питательной массы для личинок.
Энергетическая ценность цветочных масел значительно превышает таковую у сахаров, что делает их идеальным ресурсом для развития потомства. У вида Centris vittata (Lepeletier) наблюдается использование как нектара, так и масел в личиночном корме, согласно данным Pereira and Garofalo (1996). Обширный обзор этой темы был представлен Buchmann (1987). Такие адаптации демонстрируют узкую специализацию в тропических и субтропических экосистемах.
Пчелы рода Macropis из семейства Melittidae собирают масло с растений рода Lysimachia (Primulaceae). Этот ресурс используется ими не только в пищу, но и для гидроизоляции ячеек расплода, как установили Cane et al. (1983). Взрослые особи практически не употребляют масло внутрь, сохраняя зависимость от нектара как основного источника энергии. Поскольку масличные цветы часто лишены нектара, такие пчелы вынуждены посещать другие растения для собственного питания.
Для сбора масла насекомые используют специализированные подушечки, щетки или гребни из уплощенных сет (щетинок) (рис. 6-3, 108-3a). Эти морфологические приспособления позволяют эффективно впитывать или соскребать вязкий секрет. Исследования Vogel (1966–1990), а также Neff and Simpson (1981) иллюстрируют разнообразие этих структур. Они встречаются у триб Centridini, Ctenoplectrini, Tapinotaspidini и Tetrapediini в семействе Apidae.
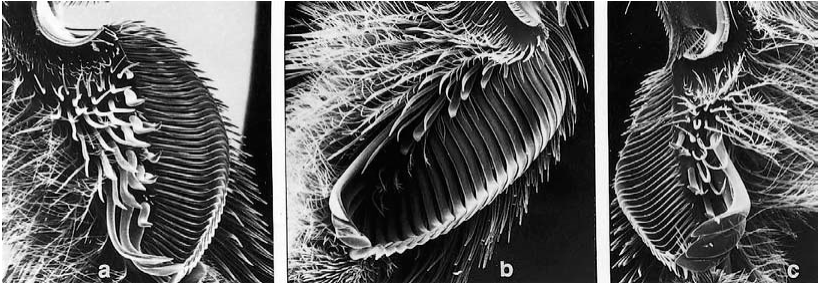
Рисунок 6-3: Вид с вентральной стороны на базитарсусы самок: (a) Centris (Ptilotopus) sp.; (b) C. (Paracentris) близ tricolor Friese; (c) C. (Heterocentris) trigonoides Lepeletier, демонстрирующие гребни из щетинок для сбора масла (по Neff and Simpson, 1981).
Использование масел развивалось независимо в разных группах пчел и семействах растений, что подтверждает Vogel (1988). В Голарктике и Старом Свете наблюдается строгая привязанность сборщиков масла к определенным родам растений. Например, Ctenoplectra связана с Momordica и Thladiantha, а Rediviva — преимущественно с Diascia. В Неотропиках специализация менее жесткая, и пчелы могут посещать различные масличные культуры.
Сбор и транспортировка пыльцы. Для большинства пчел пыльца служит основным источником белка, липидов и витаминов. Она необходима для выкармливания личинок и критически важна для полового созревания взрослых самок. Изучение тысяч социальных пчел подсемейства Halictinae (в основном Lasioglossum, подрод Dialictus) подтвердило эту зависимость. Пыльца в зобике часто обнаруживается у молодых особей с развивающимися яичниками и у активно откладывающих яйца маток.
У старых рабочих особей с редуцированными яичниками потребление пыльцы минимально или отсутствует. Однако молодые рабочие пчелы активно питаются ею для стимуляции развития экзокринных желез, как отмечают Cruz-Landim and Serrao (1994). Эти железы вырабатывают маточное молочко или другие секреты для кормления личинок. Таким образом, пыльца является фундаментом социальной структуры и репродуктивного успеха колонии.
Пыльца прикрепляется к телу пчелы благодаря шипикам, липкости поверхности или электростатическим зарядам. При перелете пчела накапливает положительный заряд, который притягивает отрицательно заряженные пыльцевые зерна. Некоторые виды транспортируют пыльцу в сухом виде, удерживая её между волосками. Другие же, например corbiculate Apinae, увлажняют её нектаром, формируя плотные обножки на задних ногах.
Сборщики масел могут использовать цветочные масла для фиксации пыльцы на скопе (scopa). Часть ресурса может переноситься в зобике для личного питания или последующей отрыжки в ячейки гнезда. Пчелы подсемейств Hylaeinae и Euryglossinae полностью лишены внешней скопы и переносят всю пыльцу внутри тела. Это считается примитивной или специализированной чертой в зависимости от филогенетической линии.
Thorp (1979) предоставил фундаментальный обзор структурных и поведенческих адаптаций к сбору пыльцы. Особое внимание исследователей привлекло строение волосков, предназначенных для манипулирования микроскопическими зернами. Работы Braue (1913), Roberts and Vallespir (1978) и Muller (1996d) углубили понимание этих процессов. Разнообразие морфологии скопы напрямую коррелирует с типом и размером пыльцы предпочитаемых растений.
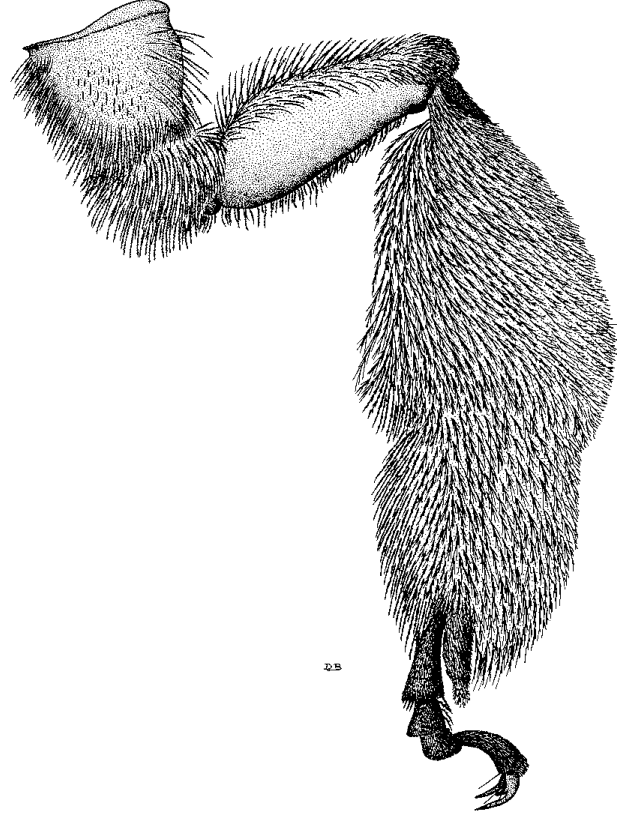
Рисунок 6-4: Задняя нога самки Svastra obliqua (Say), пчелы из трибы Eucerini, демонстрирующая скопу для транспортировки сухой пыльцы на голени и базитарсусе. Оголенная область на нижней внешней поверхности бедра образует бедренную корбикулу, характерную для многих короткоязыких пчел (рисунок D. J. Brothers).
Механизмы манипуляции пыльцой и морфология ног. Самки пчел используют модифицированные чистящие движения (grooming movements) для обработки собранной пыльцы. Сначала пыльца извлекается из пыльников передними лапками или оседает на теле при контакте с цветком. Затем передние ноги пропускаются через ротовой аппарат для питания или через средние ноги. Средние ноги, оснащенные феморальными и тибиальными щетками, счищают ресурс и передают его назад.
Конечным пунктом транспортировки являются скопы на задних ногах или, у семейства Megachilidae, метасомальные скопы на брюшке. Пыльца, осевшая на теле, тщательно вычесывается и перемещается назад в процессе полета или отдыха. Jander (1976) и Thorp (1979) описали эти движения как таксономически значимые признаки. У корбикулятных пчел (Apini, Bombini, Euglossini, Meliponini) этот процесс достигает наивысшей сложности.
На задних голенях этих пчел скопы редуцированы до состояния пыльцевых корзинок или корбикул (рис. 100-2, 118-11). Это позволяет переносить большие объемы увлажненной нектаром пыльцы с минимальными потерями. Формирование обножки требует точной координации всех пар конечностей. Такие адаптации позволяют эффективно эксплуатировать массово цветущие растения в различных климатических зонах.
Стратегии питания: Полилектия, Олиголектия и Монолектия. Полилектичные пчелы, такие как медоносная пчела Apis mellifera (Linnaeus), собирают пыльцу с широкого спектра неродственных растений. Это обеспечивает гибкость выживания в меняющихся условиях среды. В противовес им, олиголектичные виды специализируются на конкретных таксонах растений. Широкие олиголекты посещают несколько видов одного семейства, тогда как узкие олиголекты ограничены лишь несколькими близкородственными видами.
Границы между этими категориями часто размыты и представляют собой эволюционный континуум. Muller (1996b) предложил количественные критерии: олиголекты собирают более 95% пыльцы из одного семейства или трибы. Виды с «сильным предпочтением» собирают от 70% до 94% специфичной пыльцы. Остальные считаются полилектами, хотя обилие конкретных растений в регионе может искажать эти статистические данные.
Термин монолектия используется редко и применим к видам, собирающим пыльцу только с одного вида растения. Это часто происходит в условиях отсутствия родственных видов растений в ареале пчелы. Например, около 22 видов пчел в США собирают пыльцу исключительно с Larrea divaricata, согласно Hurd and Linsley (1975). Хотя их называют олиголектами, отсутствие других видов Larrea делает их фактически монолектичными в данной местности.
Интересным примером является Anthemurgus passiflorae (Robertson), специализирующаяся на Passiflora lutea. Эта пчела игнорирует другие виды Passiflora, даже если они произрастают рядом, из-за различий в размере и цвете цветков. Это указывает на глубокую поведенческую и, возможно, сенсорную адаптацию. Такие узкие связи делают насекомых крайне уязвимыми к изменениям флористического состава среды.
Специализация и коэволюция: Пчелы и их кормовые растения. Многие олиголектичные таксоны состоят из родственных видов, разделяющих пристрастие к одним и тем же растениям. Все виды рода Systropha (Rophitinae) связаны с пыльцой Convolvulus. Род Macropis неизменно использует ресурсы Lysimachia. Группа Proteriades из рода Hoplitis специализируется на Cryptantha, хотя иногда посещает другие цветы ради нектара, что приводит к случайному сбору примесей пыльцы.
Несмотря на зависимость пчел от растений, последние редко зависят исключительно от своих специализированных опылителей. Растения часто успешно размножаются за пределами ареала узких специалистов благодаря помощи полилектичных насекомых. Michener (1979a) привел множество примеров, подтверждающих этот асимметричный характер связей. Пчелы адаптируются к морфологии и химии растений, но растения реже эволюционируют под конкретный вид олиголекта.
Однако существуют явные признаки морфологической адаптации пчел к специфической пыльце. Пчелы Tetralonia malvae и Diadasia имеют редкие, грубо разветвленные волоски скопы для сбора крупной пыльцы Malvaceae и Cactaceae. У некоторых видов развились крючковидные волоски на ротовых частях или передних лапках. Эти структуры позволяют извлекать пыльцу из глубоких и узких венчиков, недоступных для других насекомых.
Примеры таких адаптаций включают Andrena osmioides и европейских пчел Colletes nasutus и Andrena nasuta. Все они являются специалистами по семейству Boraginaceae, как указал Muller (1995). Пыльца семейства Onagraceae, связанная нитями висцина, требует особого строения скопы из простых редких щетинок. Такие особенности характерны для подродов Svastra (Anthedonia) и Lasioglossum (Sphecodogastra).
Узкоспециализированные адаптации к Lamiaceae и Scrophulariaceae. Особая морфологическая черта, возникшая конвергентно, помогает пчелам собирать пыльцу с растений семейств Lamiaceae и Scrophulariaceae. Речь идет о лицевом волосяном покрове, состоящем из коротких, жестких волосков с утолщенным основанием и крючковидными кончиками. Эти волоски обычно располагаются на клипеусе (clypeus), но у рода Rophites они находятся на фронсе (frons). Такая специализация позволяет эффективно взаимодействовать с двугубыми цветками.
В случаях высокой специализации лицо пчелы становится более плоским по сравнению с родственными видами. Muller (1996a) изучил европейские примеры и подтвердил их олиголектию на губоцветных. Пчелы трутся лицом о пыльники, после чего счищают пыльцу передними лапками для переноса в скопы. Это уникальный пример того, как поведение и морфология объединяются для эксплуатации сложных флоральных структур.
Подобные модификации лица обнаружены у представителей родов Caupolicana, Andrena, Rophites, Anthidium, Trachusa, Osmia и Megachile. Также они встречаются у высокоорганизованных пчел Anthophora, Amegilla, Habropoda и Tetraloniella. Эти адаптации подчеркивают пластичность эволюции пчелиных в ответ на разнообразие растительного мира. Изучение таких связей критически важно для сохранения биоразнообразия и устойчивости агроэкосистем.
Сведения об авторах и источниках:
Авторы: Charles D. Michener
Источник: Пчелы всего мира
Данные публикации будут полезны студентам биологических специальностей, специализирующимся на энтомологии, аспирантам и исследователям в области эволюционной биологии, этологии и генетики насекомых, а также пчеловодам и всем, кто интересуется репродуктивными стратегиями и биологией развития общественных и одиночных пчел.
Дата добавления: 2026-03-15; просмотров: 232;
Поиск по сайту
Узнать еще
- Анатомическая терминология и морфология пчел: строение головы и диагностические признаки
- Вибрационное опыление и эволюция пчел: механизмы специализации и экологии
- Гаплодиплоидная система определения пола и репродуктивная биология пчел: морфология, развитие и эволюционные стратегии
- Гербаризация водных растений, оформление коллекций. Сбор гербария, оснащение
- Индустриализация строительства и внедрение сборного железобетона
- Клептопаразитизм у пчел: биологические особенности и эволюционные стратегии
- Коллемболы (ногохвостки): экология, адаптации и роль в экосистемах как почвенных инженеров
- Маслосборное корыто мотора

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
