Клептопаразитизм у пчел: биологические особенности и эволюционные стратегии
Введение в феномен клептопаразитизма. В мире пчел существует особая экологическая группа организмов, известная как клептопаразиты, которые составляют значительную часть общего разнообразия этих насекомых. Клептопаразитизм представляет собой форму трофического паразитизма, при котором взрослая особь проникает в гнездо пчелы-хозяина и откладывает собственное яйцо в подготовленную ячейку с пищевыми запасами. В подавляющем большинстве случаев взрослый паразит после завершения яйцекладки покидает гнездо, хотя некоторые виды, например представители подрода Hoplostelis s. str., могут изгонять хозяина и оставаться в захваченном гнезде. Вылупившаяся личинка паразита потребляет кормовые запасы, первоначально предназначенные для развития личинки хозяина, что послужило основанием для образного названия таких насекомых — «пчелы-кукушки». Значительно реже в природе наблюдается внутривидовой клептопаразитизм (Field, 1992), когда пчела вскрывает ячейку другой особи собственного вида и замещает отложенное яйцо своим.
Большинство клептопаразитических видов относятся к облигатно паразитирующим таксонам различного ранга — от отдельных родов до целых подсемейств. Хозяевами преимущественно выступают одиночные пчелы, однако некоторые представители общественных галиктин также могут служить объектами паразитирования. В таблице 8-2 представлен систематический обзор таксонов клептопаразитов, причем некоторые роды идентифицированы как паразитические исключительно на основании морфологических признаков — редукции или полного отсутствия структур, предназначенных для манипуляции и транспортировки пыльцы, особенно скопы у самок, а также их предположительной связи с определенными хозяевами. В упомянутой таблице такие формы обозначены буквой «р» (вероятные), а ценная информация по данной группе организмов была опубликована Грутте (Grutte, 1935).


Таблица 8-2. Систематика и особенности клептопаразитических пчел. Примечания: (p) Вероятный клептопаразит (см. раздел 8c); (n) Обнаружен в гнездах хозяев или выращен из них, но механизм уничтожения яиц или личинок неизвестен; (ad) Яйцо или личинка хозяина уничтожаются взрослой самкой паразита (предполагается при отсутствии клеток с одновременным содержанием яиц хозяина и паразита); (lo) Гибель яйца или молодой личинки хозяина происходит от активной личинки паразита с обычной морфологией; (lm) Уничтожение обеспечивается специализированной молодой личинкой со склеротизированной головой и серповидными мандибулами. Цифры в скобках указывают на вероятное количество независимых случаев возникновения клептопаразитизма. Звездочка (*) обозначает таксоны, все виды которых являются паразитическими. Социальные паразиты в список не включены (см. таблицу 8-1).
Поведенческие механизмы яйцекладки и стратегии паразитирования. Самки преимущественно клептопаразитических родов, такие как представители рода Sphecodes (триба Halictini), демонстрируют активную стратегию уничтожения потомства хозяина. Они проникают в ячейку, удаляют яйцо хозяина и замещают его собственным, что подтверждается отсутствием ячеек с двумя или более яйцами. Яйца Sphecodes морфологически практически не отличаются от яиц других галиктиновых пчел, однако совокупность поведенческих признаков — разрушение яйца хозяина в сочетании с типичной структурой молодых личинок паразита — позволяет уверенно идентифицировать этот механизм. В экспериментальном исследовании Sick и др. (1994) было задокументировано, как самка сфекодеса в наблюдательном гнезде Las ioglossum (Evylaeus) malachurum (Kirby) расширяет отверстие ячейки хозяина, проникает внутрь головой вперед примерно на две минуты (предположительно уничтожая яйцо), затем выходит, разворачивается, входит задом наперед, проводит около пяти минут (вероятно, откладывая яйцо), после чего запечатывает ячейку и на следующий день закрывает вход в гнездо. Роды, для которых установлен данный механизм, обозначены в таблице 8-2 как «ad» (уничтожение взрослой особью).
Однако самки большинства клептопаразитов не уничтожают яйцо или личинку хозяина непосредственно. В таких случаях существуют две основные стратегии откладки яиц. Первая стратегия заключается в помещении яйца в стенку незапечатанной ячейки в отсутствие хозяина, где оно скрыто от обнаружения. Вторая стратегия предполагает откладку яйца в уже готовую и запечатанную ячейку через специальное отверстие, которое самка-паразит создает и впоследствии аккуратно заделывает. Яйца, скрываемые в стенках ячеек, отличаются необычно малыми размерами по сравнению с яйцами непаразитических пчел сходного размера и демонстрируют значительное морфологическое разнообразие, часто существенно отклоняясь от типичных пчелиных яиц с мягким хорионом. Специализированные яйца характерны для всех представителей подсемейства Nomadinae, а также для трибы Protepeolini в составе Apinae и рода Coelioxys в составе Megachilini. Эти яйца внедряются во внутреннюю поверхность стенок ячейки хозяина или иным образом прячутся до её запечатывания. Напротив, яйца, откладываемые в готовые закрытые ячейки (характерные для Melectini, Ericrocidini и других), имеют обычные размеры и форму, напоминая яйца сфекодесов.
Морфологические адаптации яиц и личинок. В подсемействе Nomadinae наблюдается исключительное разнообразие не только морфологии яиц, но и способов их интродукции в стенки ячеек хозяина (Rozen, 1991a, 1992a). Некоторые виды, например Doeringiella, полностью закапывают яйца перпендикулярно стенке ячейки. Другие, как Nomada, откладывают яйца частично, оставляя один конец выступающим (см. Радченко, 1981). Третьи, например Oreopasites, могут откладывать яйца по два в одной ячейке. Некоторые виды размещают яйца почти параллельно поверхности стенки, обнажая одну сторону, которая со временем твердеет и приобретает шероховатую текстуру, контрастирующую с типично мягким хорионом (описано для Biastini Розеном, Ройг-Альсиной и Александром, 1997). Вид Epeolus, откладывающий яйца в ячейки Colletes, состоящие из двух целлофано-подобных слоев, помещает свое яйцо между этими слоями, оставляя передний конец открытым. Самки кочевых родов обладают характерными морфологическими структурами, особенно на S6 (шестом стерните брюшка), предположительно адаптированными для их специфических методов яйцекладки (рис. 8-10f, 89-2, 93-3c, d).
В отличие от непаразитарных пчел, обычно откладывающих по одному яйцу в ячейку, многие клептопаразиты, такие как Nomada и Coelioxys (представляющие разные семейства), часто помещают от двух до нескольких яиц в зараженную ячейку. Вылупившиеся личинки впоследствии уничтожают не только яйцо или личинку хозяина, но и конкурентов своего вида, пока в ячейке не остается только один выживший. Некоторые личинки клептопаразитов проявляют активность и способность уничтожать яйца или личинок хозяина, используя обычные по размеру, но острые мандибулы (например, личинки Stelis, обозначенные как «lo» — личинка с обычным строением — в таблице 8-2). Как правило, эту функцию выполняют молодые личинки, хотя Розен (Rozen, 1987a) указал, что даже личинки Stelis на последней стадии могут обладать модификациями для уничтожения хозяина.
Напротив, молодые личинки Coelioxys и паразитических Apidae обладают крупными, обычно прогнатическими, сильно склеротизированными головами и серповидными мандибулами, предназначенными исключительно для уничтожения яиц хозяина или молодых личинок. Термин «прогнатический» означает направленную вперед голову и ротовой аппарат, в отличие от более или менее направленной вниз (гипогнатической) головы, характерной для личинок других пчел. У клептопаразитических Apidae личинки первой стадии специализируются на уничтожении хозяина или конкурентов, тогда как у Coelioxys (Megachilidae) наиболее крупные мандибулы появляются у личинок второй или третьей стадии (рис. 82-5c, d). Розен (1991a) детально описал и проиллюстрировал известных личинок первой стадии паразитических триб Apinae (исключая паразитов Euglossini) и сравнил их с личинками первой стадии Nomadinae. Только у трибы Rhathymini среди паразитических Apinae головная капсула личинки первой стадии не полностью склеротизирована, сохраняя более или менее сферическую и гипогнатическую форму. Таким образом, морфология и предположительно поведение молодых личинок демонстрируют конвергентную эволюцию между различными филогенетическими группами, независимо происходящими от непаразитарных предков.
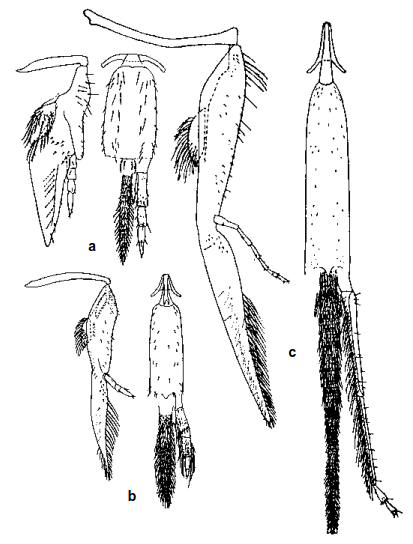
Рисунок 8-1. Сравнение ротовых структур пчел-аллодапинов: социальные паразиты Eucondylops reducta Michener (а) и Nasutapis straussorum Michener (b) демонстрируют редукцию размеров хоботка и изменение сегментации щупиков по сравнению с непаразитическим видом Allodapula melanopus (Cameron) (c). Все рисунки выполнены в одинаковом масштабе. Модифицировано по Michener, 1970.
Происхождение и эволюция клептопаразитизма. Цифры, следующие за названиями подсемейств в таблице 8-2, указывают на предполагаемое количество независимых случаев возникновения клептопаразитизма в этих группах. Однако эти оценки могут быть завышены, поскольку филогенетический анализ остается неполным или проблематичным. Например, роды Radoszkowskiana и Coelioxys могут не представлять собой отдельные линии возникновения паразитизма, а Afrostelis, Larinostelis, Stelis и, возможно, Euaspis могли произойти от одного общего паразитического предка. Трибы Protepeolini и Isepeolini и даже, возможно, Osirini могут представлять собой базальные ветви Nomadinae и, следовательно, иметь общего паразитического предка с этим подсемейством. У Melectini, Ericrocidini и Rhathymini также предположительно был общий паразитический предок. Таким образом, общее количество независимых источников возникновения клептопаразитизма может быть меньше 29, указанных в таблице. Александер (Alexander, 1990) в своем консервативном списке, исключающем сомнительные элементы, такие как Hylaeinae и Osmiini, перечислил 17 достоверных случаев возникновения паразитизма; Halictus (Paraseladonia) в то время оставался непризнанным.
Предполагаемый клептопаразитизм представителей семейства Colletidae требует дополнительного подтверждения. Перкинс (Perkins, 1899) сообщил о пяти видах гавайских Hylaeus (Nesoprosopis) как о паразитах других представителей того же подрода. Он обнаружил этих насекомых в гнездах предполагаемых хозяев и идентифицировал их по редуцированным волоскам на передних лапках самок, служащим для сбора пыльцы, однако не предоставил убедительных данных, подтверждающих его выводы. Тем не менее, учитывая общую надежность его научных работ, эта информация заслуживает внимания, хотя вопрос остается открытым для дальнейших исследований.
Несмотря на значительное таксономическое разнообразие групп пчел, у которых развивается клептопаразитизм, многие крупные, древние и широко распространенные группы не выработали этой стратегии. За исключением упомянутых выше неясных случаев, Colletidae представляют собой такую группу. Паразитические виды остаются неизвестными среди Andrenidae, Nomiinae и Rophitinae (в составе Halictidae), Melittidae, а также различных крупных групп Apidae, таких как Ceratinini, Xylocopini и Eucerini.
Правило Эмери и специфичность к хозяевам. Хозяевами пчел-клептопаразитов всегда выступают другие пчелы. Правило Эмери (см. Wilson, 1971) для паразитических перепончатокрылых насекомых гласит, что паразиты обычно нападают на своих близких родственников. Пчелы-клептопаразиты, сходные со своими непаразитическими родственниками и принадлежащие к одному роду, трибе или подсемейству, действительно паразитируют преимущественно на членах этих же таксонов. Так, паразитические Halictini в основном поражают других Halictini, хотя, как отмечалось в отношении рода Sphecodes (раздел 64), несколько видов этого рода могут паразитировать на других галиктидах и даже на пчелах из иных семейств. Паразитические эуглоссины паразитируют исключительно на других эуглоссинах. Паразитические мегахилины нападают на других представителей своего семейства, за некоторыми исключениями: отдельные виды многочисленного рода Coelioxys атакуют Anthophora, Centris, Euglossa или, по литературным данным, Tetralonia (Бишофф, 1927: 398), а Hoplostelis s. str. паразитирует на Euglossini. Насколько известно, все паразитические трибы Apinae поражают другие трибы того же подсемейства, за исключением Isepeolini, которая паразитирует на Colletinae. Как отмечалось ранее, исепеолини могли представлять собой обособленную ветвь кочевых пчел.
Интересной особенностью пчел-клептопаразитов является значительная изменчивость размеров тела. Иногда наблюдаются два отчетливых размерных класса в пределах одного вида, что традиционно объяснялось паразитированием на видах-хозяевах разного размера. Однако это предположение редко подвергалось экспериментальной проверке. Существуют три основные гипотезы, объясняющие данное явление: (1) существование криптических видов, каждый из которых специализируется на определенном виде-хозяине; (2) наличие специализированных «рас», адаптированных к конкретным хозяевам; (3) прямое влияние количества доступной пищи на рост и развитие личинок паразитического вида. Исследование Coelioxys funeraria Smith, паразитирующего на двух видах мегахил разного размера в Мичигане, подтверждает третье объяснение. На основе анализа 41 локуса генетические различия между крупными и мелкими образцами Coelioxys, собранными от двух разных хозяев, полностью объяснялись статистической ошибкой выборки (Packer et al., 1995), что свидетельствует о существовании единой панмиктической популяции паразита, атакующей оба вида мегахил.

Рисунок 8-2. Сравнение морфологии самок Halictini: а — клептопаразитический вид Sphecodes monilicornis (Кирби) с характерной грубой пунктуацией и сильными дорсолатеральными углами переднеспинки; б — непаразитический вид Lasioglossum malachurum (Кирби). На левой половине каждого рисунка волосяной покров удален. Рисунок Д. Дж. Бразерса из Michener, 1978b.
Номадины как крупнейшая группа клептопаразитов. Крупнейшей группой клептопаразитов является подсемейство Nomadinae, которое, несмотря на принадлежность к семейству Apidae, настолько существенно отличается от своих непаразитических родственников, что их ближайшие филогенетические связи остаются не вполне ясными. Роды Nomadinae часто демонстрируют значительную специфичность к хозяевам. Например, Epeolus паразитирует преимущественно на Colletes, а Doeringiella — в основном на представителях трибы Eucerini. Однако в целом подсемейство паразитирует на чрезвычайно широком спектре пчел, включая семейства Colletidae (Colletinae, Diphaglossinae), Andrenidae (все основные подсемейства), Halictidae (все подсемейства), Melittidae (Melittinae, Dasypodainae) и Apidae (Anthophorini, Eucerini, Exomalopsini, Tapinotaspidini).
Примечательной особенностью биологии различных видов кочевок является химическая мимикрия. Исследования показали, что головные выделения самцов по химическому составу напоминают летучие компоненты секрета железы Дюфура самок видов-хозяев (Andrena или Melitta) (Tengo and Bergstrom, 1976, 1977). Виды Nomada характеризуются выраженной специфичностью к хозяевам, и для каждой изученной пары «хозяин-паразит» обнаружено отчетливое химическое сходство. Секрет железы Дюфура используется непаразитическими пчелами для гидроизоляции стенок выводковых ячеек, и его запах, предположительно, служит видоспецифичным маркером гнезда. Выделения самцов андрен сходны с выделениями самок своего вида, тогда как выделения самцов и самок кочевок существенно различаются между собой. По-видимому, в процессе эволюции возникла миметическая связь между запахом хозяина (или его гнезда) и запахом самца-паразита. Функциональное значение этого явления остается предметом дискуссий: возможно, самка кочевки получает от самца во время спаривания необходимый запах для идентификации гнезда хозяина при яйцекладке, или же этот запах помогает ей проникать в гнездо, минуя агрессивную реакцию охраняющих его хозяев. Однако ни одно из существующих объяснений не является полностью удовлетворительным и требует дальнейших исследований.
Эволюционные ограничения и распространение клептопаразитизма. Анализ распространения клептопаразитизма среди пчел выявляет любопытную закономерность: несмотря на очевидные эволюционные преимущества этой стратегии, многие крупные и эволюционно успешные группы остаются свободными от облигатных паразитов. Семейство Colletidae, за исключением неподтвержденных данных по гавайским Hylaeus, полностью лишено специализированных клептопаразитов. Аналогичная ситуация наблюдается в семействе Andrenidae, подсемействах Nomiinae и Rophitinae (в составе Halictidae), семействе Melittidae, а также в таких обширных группах Apidae, как Ceratinini, Xylocopini и Eucerini. Причины отсутствия клептопаразитов в этих таксонах могут быть связаны с особенностями гнездовой биологии, эффективностью защитного поведения хозяев либо с историческими факторами эволюции сообществ.
Особый интерес представляет вопрос о множественном происхождении клептопаразитизма в эволюции пчел. Данные таблицы 8-2, указывающие на 29 независимых событий возникновения паразитизма, вероятно, отражают максимальную оценку. Филогенетический анализ позволяет предполагать, что некоторые случаи паразитизма в близкородственных группах могут иметь единое происхождение. Так, в пределах трибы Megachilini возможно родство паразитических форм Radoszkowskiana, Coelioxys, Afrostelis, Larinostelis, Stelis и Euaspis, восходящее к общему предку. В подсемействе Apinae трибы Protepeolini, Isepeolini и Osirini могут представлять собой базальные ветви Nomadinae, что предполагает их происхождение от общего паразитического предка с этим подсемейством. Трибы Melectini, Ericrocidini и Rhathymini также, вероятно, имеют общего предка. Таким образом, реальное количество независимых случаев возникновения клептопаразитизма может быть существенно меньше и составлять, по консервативным оценкам Александера (Alexander, 1990), около 17 событий.
Понимание эволюционных путей возникновения и диверсификации клептопаразитических пчел имеет фундаментальное значение для изучения коэволюционных процессов в сообществах насекомых-опылителей. Дальнейшие филогенетические исследования с применением молекулярно-генетических методов позволят уточнить родственные связи между паразитическими и непаразитическими группами и пролить свет на механизмы, способствующие или препятствующие переходу к паразитическому образу жизни у этих важнейших компонентов наземных экосистем.
Сведения об авторах и источниках:
Авторы: Charles D. Michener
Источник: Пчелы всего мира
Данные публикации будут полезны студентам биологических специальностей, специализирующимся на энтомологии, аспирантам и исследователям в области эволюционной биологии, этологии и генетики насекомых, а также пчеловодам и всем, кто интересуется репродуктивными стратегиями и биологией развития общественных и одиночных пчел.
Дата добавления: 2026-03-15; просмотров: 201;
Поиск по сайту
Узнать еще
- Активные и пассивные операции Сберегательного банка России: структура и особенности
- Анатомическая терминология и морфология пчел: строение головы и диагностические признаки
- Анатомические и физиологические особенности птиц. Перьевой покров
- Анатомия земноводных: методика вскрытия и особенности внутреннего строения на примере травяной лягушки (Rana temporaria L.)
- Анатомия костистых рыб: методика вскрытия и особенности внутреннего строения на примере щуки (Esox lucius L.)
- Анатомия пресмыкающихся: вскрытие кавказской агамы (Agama caucasica) и особенности строения рептилий
- Анатомия птицы: систематическое положение, внешнее строение и особенности вскрытия сизого голубя
- Анатомия речной миноги: особенности строения и систематика

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
