Явление адаптации. Значение рецептивного поля в координации рефлекса
Во всех рецепторах частота импульсов падает при длительном раздражении; как говорят, рецептор приспосабливается к раздражению, адаптируется. Но скорость адаптации не одинакова у разных рецепторов. Дольше всего разряжаются импульсами возбуждения мышечные рецепторы—до 10 сек., в то время как в рецепторах кожи разряд затухает а течение 1 сек.
Если рецептор приспосабливается медленно, то ритм возбуждения тем выше, чем сильнее раздражение.. Если же рецептар приспосабливается быстро, то меняется главным образом продолжительность разряда: более сильное раздражение дает более длительный ряд импульсов возбуждения. Собственно говоря, от рецепторов глубокой чувствительности, сигнализирующих о положении тела, непрерывно идут импульсы.
Это наиболее медленно адаптирующиеся рецепторы. Малейшее передвижение конечностей сразу вызывает сильный разряд импульсов в рецепторах глубокой чувствительности, который продолжается, пока движется конечность. Когда двигательный эффект прекращается, поток импульсов значительно ослабевает (Эдриан, 10).
Явление адаптации подробно было исследовано на мышечных рецепторах. Оказалось, что при растяжениях мышцы адаптация происходит тем скорее, чем меньше нагрузка (рис. 28). После прекращения: растяжения адаптация не исчезает сейчас же, а остается еще некоторое время. Например, при нагрузке 2—5 г и разгрузке через 5 сек. адаптация проходит в течение 10 сек. Повторение нагрузки ускоряет адаптацию, т. е. начальный .высокий ритм падает до более низкого уровня скорее при повторении нагрузки.
Состояние адаптации не ставится в связь с утомлением; утомление рецептора тоже имеет место, но адаптация в смысле быстрого падения начального ритма не является следствием утомления рецептора. Адаптация исчезает после растяжения в течение секунд, в то время как утомление проходит много, позднее, спустя несколько минут (Бронк, 9; Мэтьюс, 8).
По данным Эдриана, наибольшей адаптацией обладают тактильные рецепторы. Степень их адаптации близка к адаптации нервного волокна. Однако в различных тактильных рецепторах одной и той же области кожи адаптация может быть выражена в различной степени (Дзидзишвили, 20).
На рис. 18 показано, что при слабом тактильном раздражении кожи лягушки (прикосновение волоском) может возникать импульсация различной продолжительности: при раздражении некоторых рецепторов возникают всего лишь 1—2 импульса, при раздражении же других тактильных рецепторов возникает серия импульсов, постепенно затухающих в течение почти 1 секунды. При давлении на кожу или же при болевых раздражениях импульсация может продолжаться несколько секунд.
Основными сведениями о рецепторах мы обязаны прежде всего Эдриану (6, 10) и его сотруднику Мэтьюсу (8). Они регистрировали биоэлектрические токи возбуждения чувствительных волокон, отходящих от рецепторов. В некоторых опытах они регистрировали токи возбуждения чувствительного нерва при раздражении лишь одного рецептора, например одного паччиниева тельца или одного мышечного веретена. Этим путем Эдриан изучил тактильные и болевые кожные рецепторы, а Мэтьюс — проприоцепторы.
Относительно природы явления адаптации сначала предполагали, что рецептор обладает определенным запасом потенциальной энергии, которым он разряжается при раздражении, а затем при отдыхе вновь восстанавливает его за счет веществ, поступающих из крови (Мэтьюс, 8). Но в последнее время при ближайшем исследовании механорецепторов кожи лягушки было сделано более конкретное заключение о происхождении адаптации.
Механорецепторы кожи лягушки древовидно разветвляются в коже и оканчиваются частью в эпидермисе тончайшей сетью голых волоконец, расположенных между эпителиальными клетками, частью под эпидермисом — отдельными колбочками (рис. 16, I). Часть волоконец в эпителии раздражается легким прикосновением, а колбочки под эпидермисом—давлением, повреждением и химическими агентами. От раздражения прикосновением возникают быстрые импульсы типа Л, которые очень быстро адаптируются.
От давления же возникают импульсы типа В и С, которые адаптируются менее быстро. Если соскоблить эпителий и таким образом удалить лежащие здесь рецепторы, то механическое раздражение производит только импульсы типа С. Далее было установлено, что при механическом раздражении кожи ее эпителиальные клетки выделяют калий (Хоглэнд, 14).
В эпителиальных клетках содержится в 16 раз больше калия, чем в окружающей их лимфе и плазме. Он там присутствует большей частью в способной к диффузии форме и вымывается оттуда, если кожу положить на несколько часов в дистиллированную воду. Калий — единственное вещество, выделяющееся из кожи под влиянием механического раздражения.
На этом основании было сделано предположение, что при механическом раздражении кожи эпителиальные клетки выделяют калий, который действует на нервные волоконца и понижает их возбудимость. Вследствие этого механическое раздражение быстро перестает раздражать нервные окончания, т. е. происходит быстрая адаптация. Восстановление от адаптации получается вследствие диффузии калия и вымывания его током крови.
Что в данном случае адаптация действительно обусловлена химическим веществом, видно из следующего. Если после наступления адаптации промыть кожу нормальным раствором Рингера, то адаптация быстро проходит, В то же время промывание кожи раствором Рингера, содержащим в 15 раз больше калия, чем в норме, заметно ускоряет адаптацию. Все это свидетельствует о том, что адаптация обусловливается калием.
Можно показать, что калий выделяется эпителиальными клетками, а не самими нервными волоконцами; это видно из того, что сейчас же после длительного раздражения кожного чувствительного нерва электрическим током нервные волоконца в эпителии, несмотря на длительное возбуждение, не оказываются адаптированными: они отвечают, как обычно, на механическое раздражение кожи. Значит, от возбуждения нервных волоконец калий не выделяется, адаптация не наступает.
На основании всех этих фактов Хоглэнд (14) имел право заключить, что быстрая адаптация голых рецепторов в эпидермисе обусловливается парализующим действием калия, выделяемого из эпителиальных клеток. Калий, как известно-, в малой концентрации повышает возбудимость нервной ткани, а в большой дозе, наоборот, понижает ее. Нужно думать, что та доза калия, которая выделяется из эпителиальных клеток, достаточна для того, чтобы подействовать на нервные волоконца угнетающим образом.
Менее быструю адаптацию на давление и повреждение кожи лягушки Xоглэнд (14) считает следствием отдаленности соответственных рецепторов от эпидермиса. Он же считает, что различная инкапсуляция многих рецепторов имеет назначение защитить рецептор от действия калия, выделяемого окружающими эпителиальными клетками при их механическом раздражении.
Значение рецептивного поля в координации рефлекса. То обстоятельство, что каждый рефлекс имеет свое рецептивное поле, дает возможность изучить роль последнего в координации рефлекса. Можно взять чувствительный нерв, отходящий от рецептивного поля данного рефлекса, и раздражать его механически, химически или электрическим током. Во всех случаях рефлекс вызывается с точно таким же постоянством и с такой же координацией, как и при раздражении самого рецептивного поля.
Так происходит и в том случае, если чувствительный нерв перерезан и раздражается его центральный отрезок. Например, на спинномозговой лягушке раздражение центрального отрезка n. peroneus, который получает волокна из рецептивного поля сгибания, дает вполне координированный рефлекс сгибания. Это было доказано и на других животных. Так, например, легкое раздражение кожи на шее, на плечах и спине кошки, собаки, кролика и других животных вызывает рефлекс чесания (рис. 20).
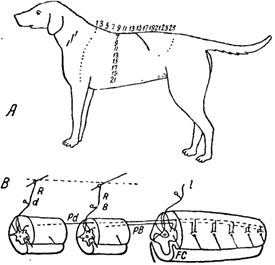
Рис. 20. Рецептивное поле рефлекса чесания на собаке (обозначено на рисунке цифрами) (А) и рефлекторная дуга чесания (В). dR, BR — чувствительные нейроны от рецептивного поля чесания; Рd, РВ — интраспинальные нейроны или внутримозговые; FC — двигательные нейроны (Шеррингтон)
Этот рефлекс состоит в притягивании задней ноги раздражаемой стороны к раздражаемому участку и в ритмических движениях ног. В это время другая задняя нога разгибается, шея и туловище поворачиваются в раздражаемую сторону. Данный сложный рефлекс вызывается также электрическим раздражением чувствительных нервов соответствующего рецептивного поля (Шеррингтон, 5).
Таким образом, рецептивное поле служит только для восприятия раздражений, в координации же рефлекса оно не участвует.
Дата добавления: 2023-04-03; просмотров: 1452;
Поиск по сайту
Узнать еще
- Вторичные и дополнительные поля
- Головной мозг. Проекционные поля коры
- Значение двигательных нейронов в координации рефлекса
- Морфология и физиология (значение, развитие, связь с другими биологическими науками)
- Назначение и устройство транспортира. Построение угла 45° с помощью транспортира
- Некоторые функциональные отделы коры. Первичные моторные и сенсорные передаточные поля
- Основные процессы центральной координации
- Рецептивное поле рефлекса и его значение

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
