Анатомическая терминология и морфология пчел: строение головы и диагностические признаки
Введение в морфологическую номенклатуру. Иллюстрации и текст настоящего раздела содержат детальные обозначения внешних структур взрослых пчел, при этом многие анатомические элементы просто маркированы на сопроводительных рисунках. Структуры, не показанные на иллюстрациях или требующие дополнительного обсуждения и пояснений, рассматриваются в текстовой части, где предпочтительные термины выделяются жирным шрифтом в местах их определения или объяснения. Основная цель данного подхода заключается в том, чтобы совокупность иллюстративного материала и текстовых описаний способствовала унификации терминологии в будущих таксономических и морфологических исследованиях, посвященных пчелам. Такой стандартизированный подход обеспечивает преемственность научной коммуникации и позволяет избежать разночтений при описании новых таксонов.
В данном разделе основное внимание уделяется признакам, которые варьируют у разных видов пчел; крайне мало терминов объясняется или иллюстрируется для обозначения структур, не предоставляющих полезных диагностических характеристик для одного или нескольких таксонов. Такой прагматичный подход позволяет сконцентрироваться на действительно значимых морфологических элементах, имеющих систематическую ценность. Описание куколок не приводится, поскольку строение этой стадии развития может быть дифференцировано от строения взрослых особей и не содержит новой терминологии, помимо уже установленной для имагинальной фазы. Таким образом, все морфологические описания ориентированы исключительно на взрослых насекомых, что соответствует практике современных таксономических ревизий.
Историческая основа и референсные исследования. Морфология медоносной пчелы (Apis mellifera Linnaeus), будучи наиболее тщательно изученной, служит эталонным материалом для исследователей, интересующихся пчелами в целом. Фундаментальный вклад в эту область внес Р.Э. Снодграсс (Snodgrass, 1956), чьи работы предоставили не только ценные сведения о морфологии пчел, но и разработали терминологию для обозначения структур, основанную на его более ранних исследованиях. Эта терминологическая система послужила основой для последующих работ по анатомии пчел и продолжает использоваться в современной литературе с соответствующими модификациями. Несколько авторов, изучающих пчел, не относящихся к роду Apis, представили собственные описания внешней морфологии с различной степенью детализации, используя терминологию Снодграсса, за исключением случаев, требующих отклонений, и вводя дополнительные термины по мере необходимости.
К числу таких значимых исследований относятся работы Ч.Д. Миченера (1944) по роду Anthophora; Урбана (1967a) по роду Thygater; Камарго, Керра и Лопеса (1967) по роду Melipona; Эйкворта (1969a) по роду Pseudaugochlora; Гербера и Акре (1969) по роду Megachile; Песенко (1983) по роду Nomioides и Брукса (1988) по трибе Anthophorini. Нет необходимости повторять здесь детали морфологии, представленные в указанных работах, однако терминология, используемая в последующих разделах данной книги, требует некоторых пояснений, следуя системе Миченера (1944) с соответствующими модификациями. Многие структуры, упоминаемые в систематизированных частях (разделы с 33 по 119), обозначены на рисунках с 10-1 по 10-15, которые носят преимущественно схематичный характер и предназначены для иллюстрации максимального количества анатомических элементов пчел.
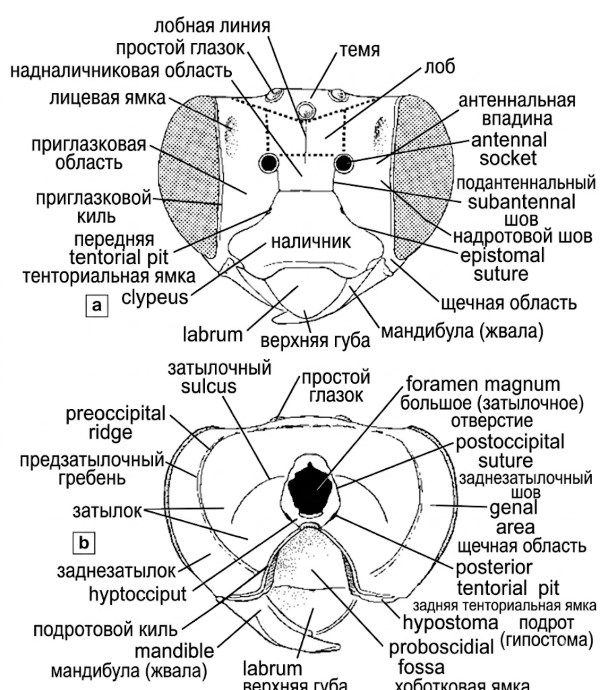
Рисунок 10-1. Диаграммы головы пчелы, показывающие основные структуры: а — вид спереди; б — вид сзади. Из работы Миченера, Макгинли и Данфорта, 1994.
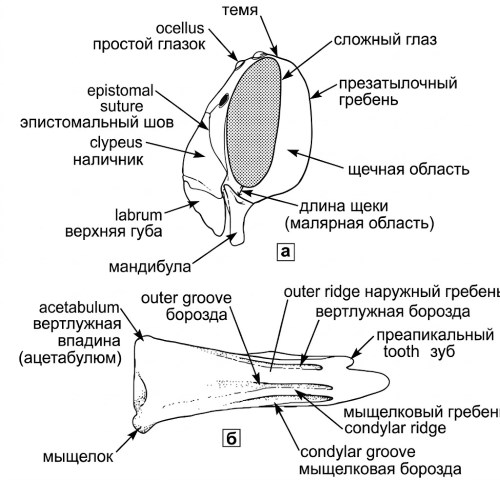
Рисунок 10-2. а — схематический вид головы пчелы сбоку; б — наружная поверхность нижней челюсти. Из работы Миченера, Макгинли и Данфорта, 1994.
Строение головы и анатомия ротового аппарата. На рисунках 10-1 и 10-2 показаны основные структуры головы, при этом в отношении номенклатуры частей нижней челюсти автор обычно следует за Миченером и Фрейзером (1978), как это проиллюстрировано на рисунке 10-2. Для простоты изложения часто используется термин преапикальные зубы по верхнему краю, а не "зубы нижней челюсти", что подразумевает верхний край мандибулы над мыщелковой впадиной, обычно заканчивающийся одним или несколькими преапикальными зубами. Рутеллум представляет собой остальную дистальную часть нижней челюсти, расположенную под головкой, и обычно формирует главный или нижнеапикальный зуб мандибулы, хотя у некоторых пчел, включая представителей рода Lithurgus и многих Xylocopinae, имеется нижний преапикальный зуб, то есть рутеллярный зуб, расположенный ниже основной вершины (рис. 76-3а, 88-9а, б).
Другие важные термины, используемые в настоящем разделе, включают мыщелковый гребень, который возникает вблизи мыщелка нижней челюсти под нижним или задним базальным углом мандибулы и простирается по направлению к ее вершине. Внешний гребень представляет собой следующий дорсальный гребень над мыщелковым гребнем на наружной поверхности нижней челюсти; эти структуры имеют критическое значение для понимания вариабельности формы мандибул у различных таксонов пчел. Изучение этих гребней позволяет выявить филогенетически значимые признаки, используемые в современной систематике.
Терминология для обозначения частей верхней губы часто вызывает путаницу из-за использования в роде Andrena термина "отросток" для обозначения базальной приподнятой пластинки, что вводит в заблуждение, поскольку эта структура не выступает так, как можно было бы ожидать от отростка. У других пчел, например, у Panurginae (см. Ruz, 1986), та же структура именуется базальной областью губы, и использование слова "отросток" в этом контексте создает дополнительную путаницу, поскольку у некоторых пчел, особенно у Halictidae, на вершине верхней губы находится совершенно иной выступ, обозначаемый здесь как апикальный отросток верхней губы. Во избежание неоднозначности рекомендуется строго разграничивать эти гомологичные структуры.
Области головы и лицевые ямки. В описательных целях поверхность головы часто подразделяется на нечетко ограниченные области, как показано пунктирными линиями на рисунке 10-1. К ним относятся две параокулярные области, расположенные вдоль глаз; надглазничная область; лобная или надантеннальная область; и вертекс (темя). Скуловая область (или скуловое пространство) находится между глазом и основанием нижней челюсти, при этом ее длина представляет собой кратчайшее расстояние от глаза до мандибулы (рис. 10-2), а ширина соответствует ширине базальной части нижней челюсти. Эти измерения часто используются в диагностических ключах для различения близких видов.
Лицевые ямки (рис. 10-1) и ямки по бокам второго тергита метасомы (рис. 10-12) представляют собой углубления, обычно пигментированные черным цветом и потому контрастно выделяющиеся на более светлом фоне основного покровов. Лицевые ямки парные, по одной в каждой параокулярной области, и располагаются преимущественно или полностью выше уровня оснований усиков, иногда достигая темени между глазками и верхними краями сложных глаз. Эти ямки, лучше развитые у самок, чем у самцов, могут быть точечными или узкими, лишенными волосков (как у многих Hylaeinae и Euryglossinae) либо представлять собой широкие, слегка вдавленные участки, тогда как у большинства Andreninae они покрыты мелкими волосками.
Шуберт и Шёнитцер (Schuberth, Schönitzer, 1993) представили детальное анатомическое описание лицевых ямок у различных таксонов, показав, что эпидермис под кутикулой состоит из секреторных клеток. Это позволяет предположить, что ямки являются испаряющими поверхностями, хотя химическая природа их секретов и точные функции остаются неизвестными. У многих пчел, лишенных отчетливых лицевых ямок, тем не менее имеются участки дифференцированной кутикулы, предположительно гомологичные ямкам. Такие участки обычно характеризуются более редкой и менее крупной пунктировкой по сравнению с соседними зонами, часто отличаются микроструктурой поверхности и цветом (как правило, черным). Таким образом, существует непрерывный ряд переходов от полного отсутствия ямок (как у Megachilidae и большинства Halictinae) до отчетливых, вдавленных ямочных структур.
Антеннальная область и сопутствующие структуры. Усики отходят от усиковых впадин, иногда называемых альвеолами; для простоты изложения вместо термина "флагелломеры" используются выражения "членики усиков" или "сегменты жгутика". Почти у всех пчел имеется субантеннальный шов, проходящий от каждой усиковой впадины до эпистомального шва. Однако если усики расположены близко к наличнику, субантеннальные швы могут отсутствовать, а у некоторых пчел (большинство Andrenidae) под каждым усиком присутствуют два субантеннальных шва, ограничивающих субантеннальную область. Эта особенность имеет важное диагностическое значение на уровне семейств.
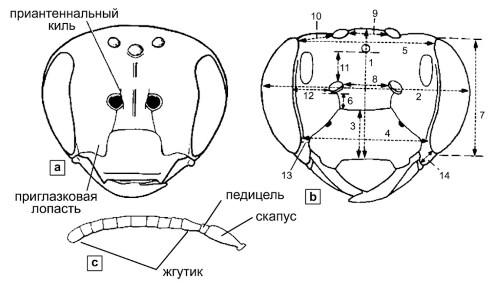
Рисунок 10-3. Схемы головы и антенн: а — схематический вид головы пчелы спереди, показывающий структуры, не изображенные на рисунке 10-1 (ни у одной известной пчелы нет одновременно параокулярных долей и прилежащих килей); б — схематический вид головы пчелы спереди с указанием способов измерений: (1) длина головы (лица), (2) ширина головы, (3) длина наличника, (4) нижнее межглазничное расстояние, (5) верхнее межглазничное расстояние, (6) клипеоантеннальное расстояние (длина субантеннального шва, если он прямой), (7) длина глаза, (8) межантеннальное или межальвеолярное расстояние, (9) межглазничное расстояние, (10) оцеллярное расстояние, (11) антенно-оцеллярное или альвеоло-оцеллярное расстояние, (12) антенно-окулярное или альвеоло-окулярное расстояние, (13) клипео-окулярное расстояние, (14) длина скуловой области; в — антенна самки пчелы. Из работы Миченера, Макгинли и Данфорта, 1994.
У некоторых пчел имеется продольный киль, расположенный непосредственно над основанием усика; он обычно приподнят чуть выше базальной части усика и часто формирует пластинку, частично прикрывающую основание усика (рис. 10-3а). Такие кили нередко называют межантеннальными или межальвеолярными, однако термин юкстаннальные кили, предложенный Миченером и Грисволдом (Michener and Griswold, 1994a), предпочтительнее, поскольку он более точно описывает их положение рядом с усиками, а не между ними. Эта терминологическая точность важна при описании тонких морфологических различий между близкими таксонами.
У некоторых пчел (например, у представителей рода Augochlora) наблюдается смещение передних тенториальных ямок вниз, что влечет за собой соответствующее изменение эпистомального шва, который становится наклонным или изогнутым, переходя на наличник с каждой стороны. В результате параокулярная область заходит на наличник, формируя структуру, называемую параокулярной долей (рис. 10-3а). Термин "орбита" часто используется для обозначения края глаза, при этом внутренняя орбита соответствует лобному или лицевому краю, а внешняя орбита — общему краю. Выражения типа "глаза сходятся книзу" обычно эквивалентны утверждению "внутренние орбиты сходятся книзу", что является важным диагностическим признаком.
Методики измерений и краниальные структуры. Описания пчел часто включают промеры или, что еще чаще, указания относительных размеров различных структур, особенно головы. На рисунке 10-3b показано, как следует производить стандартизированные измерения. При виде сверху на темя используется постокулярная касательная — воображаемая линия, проведенная по касательной к задним выпуклостям обоих глаз. Глазнично-затылочное расстояние измеряется между задним краем глаза и точкой, где темя изгибается или круто опускается к задней поверхности головы, то есть обычно до предзатылочного гребня. Эти метрические характеристики широко используются в количественной таксономии.
Затылочная область — это участок позади глаза и перед затылочным гребнем. Предзатылочный гребень окружает вогнутую заднюю поверхность головы сверху и сбоку. На этом гребне иногда выделяется гипостомальный киль, который может быть дорсальным (только за темянем), латеральным (только за глазом) или полным (охватывающим как дорсальную, так и латеральную части). Согласно Силвейре (Silveira, 1995a), дорсальная часть предзатылочного гребня у некоторых Exomalopsini на самом деле расположена на задней поверхности головы, а непосредственно за глазками находится новый поперечный гребень — постоцеллярный гребень (или киль). Это открытие демонстрирует, что даже хорошо изученные группы могут содержать ранее не описанные морфологические элементы.
Хоботковая ямка и нижнечелюстной аппарат. Хоботковая ямка (рис. 10-1) представляет собой обширное глубокое углубление на нижней стороне головы, в которое укладывается сложенный хоботок. Она ограничена с боков гипостомальным гребнем, передний конец которого изгибается латерально за основание нижней челюсти. Нижняя сторона головы, расположенная латеральнее гипостомального гребня и позади основания мандибулы, называется гипостомальной областью или, согласно Эйкворту (1969b), постгеной, хотя этот термин не охватывает всю гипостому, поскольку последняя включает также стенки и крышу хоботковой ямки. Точное описание этих структур необходимо для понимания механики работы ротового аппарата.
Парамандибулярный отросток представляет собой передний выступ гипостомы, который приближается к боковой части наличника, упирается в нее или срастается с ней; в последнем случае обеспечивается склеротическое замыкание мандибулярной впадины. Щетинки или волоски, растущие от гребня на парамандибулярном отростке и иногда продолжающиеся латерально на боковой оконечности гипостомального киля, образуют у некоторых видов Andrenidae структуру, называемую подродовой короной. Этот признак может иметь значение для внутриродовой систематики.
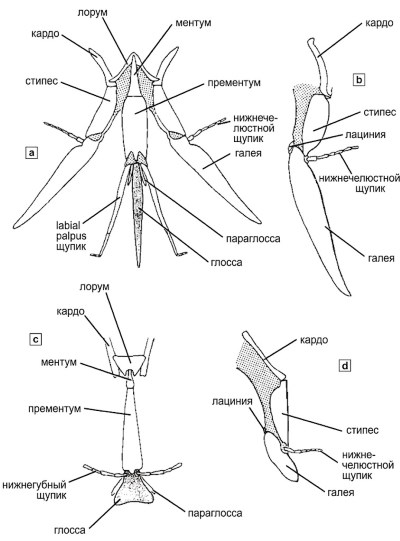
Рисунок 10-4. Диаграммы хоботков пчел: а — раздвинутый хоботок пчелы с длинным языком; б — верхняя челюсть пчелы с таким же языком; в — нижняя губа пчелы с коротким языком, в данном случае ложковидная, с изображением частей максилл у основания; г — верхняя челюсть пчелы с коротким языком. Из работы Миченера, Макгинли и Данфорта, 1994.
Максиллы и лабиум образуют сложный комплекс ротовых органов, адаптированный к потреблению жидкой пищи — нектара. Строение глоссы (язычка) варьирует от короткого и ложковидного у пчел с коротким хоботком до длинного и тонкого у специализированных опылителей цветков с глубоким венчиком. На рисунке 10-4 представлены схематические изображения хоботков пчел с различной длиной ротового аппарата, демонстрирующие соотношение лабиальных щупиков, параглосс и других элементов. Эти структуры имеют первостепенное значение для понимания трофической специализации и коэволюции пчел с цветковыми растениями.
В последующих разделах систематической части (с 33 по 119) читатель будет постоянно обращаться к терминологии, изложенной выше, что подчеркивает важность единообразного понимания морфологических структур. Представленные здесь рисунки и пояснения призваны служить справочным аппаратом при работе с таксономическими ключами и описаниями видов, обеспечивая корректную интерпретацию диагностических признаков.
Сведения об авторах и источниках:
Авторы: Charles D. Michener
Источник: Пчелы всего мира
Данные публикации будут полезны студентам биологических специальностей, специализирующимся на энтомологии, аспирантам и исследователям в области эволюционной биологии, этологии и генетики насекомых, а также пчеловодам и всем, кто интересуется репродуктивными стратегиями и биологией развития общественных и одиночных пчел.
Дата добавления: 2026-03-15; просмотров: 249;
Поиск по сайту
Узнать еще
- Анатомическая локализация координирующих аппаратов
- Анатомическое строение масличных семян
- Анатомия птицы: систематическое положение, внешнее строение и особенности вскрытия сизого голубя
- Астеносфера Земли: строение, свойства и роль в глобальной геодинамике
- Бактериофаги. Описание и строение
- Блоки зубчатых колес. Машиностроение
- Вибрационное опыление и эволюция пчел: механизмы специализации и экологии

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
