Эколого-фитоценотическая или флористическая классификация растительности?
В последние два десятилетия в отечественной геоботанической литературе усиленно пропагандируется флористическая классификация растительности (классификация по Браун-Бланке). Сторонники этого направления приводят действительные и мнимые достоинства этой классификации, одновременно подвергая жесткой критике эколого-фитоценотическую классификацию, которая называется ими доминантной. Действительно ли одна из этих классификаций значительно лучше другой? Преодолевает ли какая-то из них те объективные сложности, которые встают при классификации растительности?
На мой взгляд, приверженность тому или иному направлению в классификации не гарантирует сама по себе высокого качества классификации. Мне известны многочисленные глубокие и качественные работы представителей обоих направлений, и в то же время, как у нас, так и на Западе имеется много формально выполненных работ, дающих очень мало для понимания каких-то закономерностей растительности.
По-видимому, одной из самых привлекательных сторон флористической классификации растительности для значительного числа приверженцев этого направления является наличие строго разработанных номенклатурных правил (Кодекса фитоценотической номенклатуры). Эти правила предоставляют к тому же возможность поставить свою фамилию после названия синтаксона. Кодекс был списан с небольшими изменениями с Кодекса ботанической номенклатуры, и он плохо учитывает особенности наших объектов классификации.
В частности, очень неудачно введенное понятие о типовом описании. Я не могу представить, как можно выделить такие описания в непрерывно варьирующей растительности. По-моему, ни в одной ассоциации нельзя найти описание, которое можно было бы считать типичным. Видимо, поэтому типичные описания выделяют только в Чехии и наиболее ретивые приверженцы Кодекса в нашей стране.
Формальное следование какому-то одному критерию выделения ассоциаций и более высоких синтаксонов часто дает неудовлетворительные результаты. Неоднократно критикуемые Б. М. Миркиным классики эколого-фитоценотической классификации — В. Н. Сукачев и А. П. Шенников в своих работах по классификации растительности не были слепыми последователями классификации по доминантам. Достаточно привести работу В. Н. Сукачева (1931) «Типы леса Бузулукского бора», в которой ассоциации сосновых лесов выделены по характерным видам или по положению в рельефе, так как доминирующих видов в нижних ярусах степных сосняков очень часто нет. Сравнительно небольшое число четко охарактеризованных ассоциаций дал в своей работе «Луга Симбирской губернии» А. П. Шенников (1919).
В то же время последовательное применение доминантного подхода нередко приводило к выделению бесчисленного числа мелких ассоциаций, неповторяющихся в природе и не имеющих никаких заметных отличий по условиям среды, в которых они встречаются. По сути дела, каждая комбинация обильных видов возводилась в ранг ассоциации. Ценность таких единиц, конечно, крайне мала. Особенно заметно проявлялась эта тенденция при классификации лугов и степей, где богатый видовой состав и полидоминантность сообществ дают громадное число таких комбинаций.
Большой минус работ по флористической классификации растительности состоит в том, что все работы, выполненные вне рамок этого подхода, полностью игнорируются.
Работа начинается с чистого листа, что значительно легче, но возникает ничем не оправданный разрыв с тем, что было сделано в нашей стране ранее. А сделано было немало: разнообразие растительности России и всей территории бывшего СССР было изучено довольно детально. Много внимания уделялось географическим и ландшафтным закономерностям распределения растительности, закономерностям ее динамики и биологической продуктивности. Без учета этих работ нельзя выявить ареал любой единицы флористической классификации, сказать что-либо определенное о ее динамических тенденциях.
Вот поэтому нередко в работах по флористической классификации кроме списка дифференциальных или характерных видов ничего иного найти нельзя. Конечно, это не дефект самого флористического подхода к классификации, а наша отечественная особенность. В западноевропейских работах, где флористическая классификация существует давно, во многих работах можно найти весьма детальную информацию об ареале, среде и динамике синтаксонов. А многие из тех, кто работал и работает в рамках эколого-фитоценотического подхода, также уделяют мало внимания сравнению своих данных с материалами и выводами предшественников.
И флористическая, и эколого-фитоценотическая классификации в своей основе ие- рархичные. Строится иерархия синтаксонов, включающая значительное число уровней. В эколого-фитоценотической классификации это тип растительности, класс формаций, формация, группа ассоциаций, ассоциация. Во флористической классификации это класс, порядок, союз, ассоциация. В той и другой классификации выделяют кроме этого еще несколько промежуточных уровней. Оба подхода в этом отношении достаточно сходны.
Но любая иерархическая классификация, в общем, противоречит природе растительности. Варьирование растительности не только непрерывно, но и многомерно, идет во многих направлениях (Раменский, 1925). Как следствие этого любой синтаксон связан в отношении флористического состава с большим числом синтаксонов этого же ранга в разных экологических (по влажности, богатству и другим экологическим осям) географических, динамических рядах. Вырезанные из этого многомерного варьирования синтаксоны высокого ранга нарушают эти многомерные связи с синтаксонами того же ранга.
При характеристике многих ассоциаций геоботаники Западной Европы указывают, что в их составе имеются характерные виды самых разных союзов, порядков и классов. Это обстоятельство вызывает частые перестановки ассоциаций из одного синтаксона высокого ранга в другие. Эта проблема встает, конечно, и при доминантной классификации, но из-за ее меньшей формализованности на это мало обращают внимание. Когда-то Е. П. Матвеева (1967) разделила щучковые луга (одну формацию) по нескольким классам формаций, чем вызвала гнев оппонентов. Была нарушена строгость иерархии, но получена большая экологичность классификации.
Классификация нередко выступает как самоцель. Но это прежде всего средство хранения и поиска информации об объектах классификации. В этом отношении иерархическая классификация совершенно необходима, но сделать эту иерархию стабильной, общепринятой и безупречной вряд ли удастся. Чем большим содержанием наполняется каждая ячейка классификации, тем более полезной для геоботаников она становится и тем больше шансов, что коллеги будут относиться к ней более внимательно. Один набор доминирующих видов или два-три дифференциальных вида большого почтения у коллег не вызывают.
Во флористической классификации растительности (по Браун-Бланке) более половины ассоциаций выделены по доминированию одного вида, и этот доминант является единственным характерным видом данной ассоциации. В этом случае различия между подходами выражается только в ранге единиц: в эколого-фитоценотической классификации — это формация, во флористической — ассоциация.
Выделение ассоциаций или других единиц классификации по одному виду, доминанту или характерному виду, удобно. Такие единицы легко распознаются в поле. Их можно картировать, достаточно точно проводить границы между ними на местности, относительно легко проследить динамику. Но нужно различать признаки, по которым выделяются единицы классификации (критерии деления), и признаки, которые являются следствием деления.
Синтаксоны, полученные по признаку доминирования какого-то одного вида, только тогда заслуживают названия естественных, когда они отличаются не только по критерию деления, но имеют значительное число характеризующих их признаков (не обязательно флористических). Во флористической классификации существует союз Magnocaricion, в котором все ассоциации выделены по доминированию одного вида осоки: Caricetum gracilis, Caricetum cespitosae, Caricetum vulpinae и т.д. D. Blazkowa (1971) в статье, посвященной асс. Caricetum gracilis, писала, что в этой ассоциации нет никаких постоянных спутников Carex acuta, а встречающиеся вместе с этим доминантом виды непостоянны, довольно случайны и их нельзя использовать в качестве дифференциальных.
Отсутствие хороших дифференциальных видов в таких синтаксонах свидетельствует о том, что они недостаточно существенно отличаются друг от друга по условиям местообитания, что доминанты этих ассоциаций меняют среду вокруг себя достаточно сходным образом, вследствие чего такие синтаксоны и не имеют характерных спутников. Такие виды называют фитоценотически замещающими. Исходя из принципов, которых придерживаются сторонники флористической классификации, такие синтаксоны следовало бы объединять на уровне ассоциаций.
Другая сложность, с которой сталкивается флористическая классификация, состоит в том, что каждая ассоциация должна быть отнесена к определенному союзу, порядку и классу, а для этого в ней должны присутствовать характерные виды всех этих единиц. В действительности, такая ситуация встречается далеко не всегда. В этом случае выделяют так называемые базовые сообщества (Basalgesellschaften) класса, порядка или союза. Складывается парадоксальная ситуация, аналогичная тому, как если бы систематики растений считали, что данное растение является крестоцветным, но не может быть отнесено ни к какому роду и виду.
Все это не говорит о том, что флористический подход к классификации растительности является каким-то порочным и им ни в коем случае нельзя пользоваться. Все это отражение тех реальных сложностей, с которыми сталкиваются геоботаники при классификации непрерывно и многомерно варьирующей растительности. Флористический подход — один из возможных подходов в классификации растительности. Он имеет определенные преимущества при классификации многовидовых и полидоминантных растительных сообществ, в частности луговых и степных.
Посмотрим, как же различаются растительные ассоциации флористической классификации, по тем признакам, которые считаются в ней основными, т. е. по флористическому составу. В работе J. Jirasek (1996) приведена характеристика всех ассоциаций еловых лесов Чехии. Автор приводит для каждой из них характерные комбинации видов, т.е. видов с высоким постоянством (табл. 1). Из этой таблицы видно, что имеется группа видов, которая встречается во всех ассоциациях ельников. К ним относятся прежде всего Vaccinium myrtillus, Avenella flexuosa (Lerchenfeldia flexuosa), Calamagrostis villosa, высокое обилие которых свойственно большинству ассоциаций ельников гор Центральной Европы.
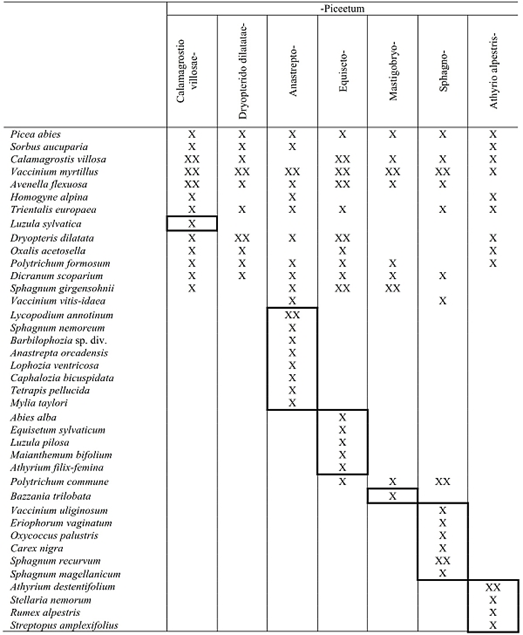
Таблица 1. Характерные комбинации видов ассоциаций еловых лесов Чехии (по: Jirasek, 1996)
Не все ассоциации содержат виды, которые входят в характерную комбинацию только одной ассоциации. В широко распространенной асс. Calamagrostio villosae-Piceetum только один такой вид — Luzula sylvatica. В асс. Mastigobryo-Piceetum — только печеночник Bazzania trilobata, а в асс. Dryopterido dilatatae-Piceetum нет ни одного такого вида. Остальные ассоциации имеют достаточно крупные группы видов, характерных для этих ассоциаций, четко отделяющие их от других ассоциаций ельников. Это прежде всего ассоциации, занимающие крайние местообитания среди ельников: асс. Sphagno-Piceetum характеризуется наличием группы болотных видов, асс. Athyrio alpestris-Piceetum встречается на почвах с большим количеством обломков известняков. Виды, характеризующие эти ассоциации, встречаются и в других ассоциациях, не включенных в эту таблицу.
Чешские геоботаники выделяют довольно узкие растительные ассоциации, объем которых сопоставим с ассоциациями, которые выделяют русские геоботаники в лесах, но как во всякой флористической классификации для них важнее дифференциальные виды, чем доминанты.
Рассмотрим еще один пример такой классификации. В работе E. Balatova-Tulackova (2000) приведена характеристика ассоциаций союза Calthion из северо-восточной Чехии. В характерные комбинации видов каждой ассоциации входит большое число видов, но только немногие из них являются специфическими для какой-то одной ассоциации (табл. 2). Из таблицы видно, что каждая ассоциация имеет небольшое число видов в характерной комбинации. К тому же, не всегда эти виды индицируют какие-то определенные экологические условия.
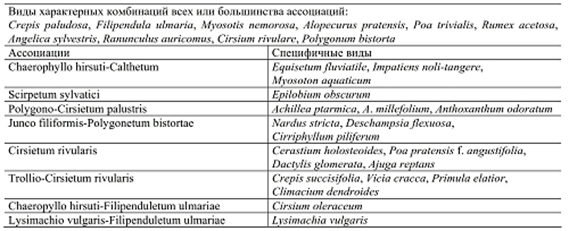
Таблица 2. Характерные комбинации видов ассоциаций союза Calthion из северо-восточной Чехии (по: Balatova-Tulackova, 2000)
Оба эти примеры взяты из работ опытных и квалифицированных геоботаников. Они приведены здесь не для критики флористического подхода, а для того, чтобы показать объективно существующие возможности получить дискретные по флористическому составу единицы классификации растительности.
Такие результаты в очень тщательно выполненных работах не дискредитируют флористический подход к классификации растительности. В работах с использованием доминантного подхода найти что-либо сопоставимое весьма сложно, так как в работах этого направления очень часто не приводятся более или менее полные флористические данные и не анализируются различия между ассоциациями по флористическому составу.
Для этой цели можно использовать только монографию И. М. Сцепановiча (2000), который использовал своеобразный подход: классифицировал луговую растительность по доминантам, но называл ассоциации по Браун-Бланке и привел в работе списки дифференциальных видов для всех ассоциаций. В табл. 3 приведены специфические дифференциальные виды пяти ассоциаций (формаций эколого-фитоценотической классификации).

Таблица 3. Специфические дифференциальные виды некоторых луговых ассоциаций Белоруссии (по: Сцепановіч, 2000)
Даже при рассмотрении только пяти ассоциаций специфические дифференциальные виды оказываются лишь в трех ассоциациях. При расширении числа ассоциаций часть из них может оказаться дифференциальными и в каких-то других ассоциациях. Festucetum rubrae дифференциальные виды характеризуют достаточно хорошо. Все они свидетельствуют о более ксерофильных условиях в этой ассоциации по сравнению с остальными. Все остальные четыре ассоциации оказываются очень плохо охарактеризованными специфичными дифференциальными видами. Почему в Brizetum mediae таким видом оказалась Fragaria vesca, а в Anthoxanthethum odoratae — Veronica officinalis? Скорее всего, это объясняется случайным выборочным варьированием, вследствие которого вид имеет в выборке более высокую встречаемость.
Все то, что говорилось выше о классификации, не имеет прямого отношения к водной растительности, где преобладают фитоценозы с явным доминированием одного вида и очень бедным видовым составом. Но почему водная растительность отличается такими свойствами? Условия среды, хотя и довольно специфические, не отличаются крайней суровостью. Об этом говорит и значительное общее число видов водных растений во многих регионах.
Большое число доминирующих видов позволяет выделить значительное число ассоциаций. Но каждая ли из них приурочена к специфическим для нее условиям среды или же несколько таких ассоциаций могут встречаться в одном и том же типе местообитаний? В том, что разные ассоциации водных растений могут встречаться в одинаковых местообитаниях, можно убедиться, анализируя литературу по водной растительности.
В статье о водной растительности Сейненского поозерья на северо-востоке Польши H. Tomaszewicz и S. Klosowski (1985) приводят 48 ассоциаций. Для каждой из них указывается тип субстрата, на котором она встречается, и диапазон глубины воды. Это серьезные факторы дифференциации водной растительности. Многие ассоциации перекрываются по этим параметрам. Конечно, этими двумя факторами не исчерпывается все, что оказывает влияние на водную растительность. Имеет значение трофность водоемов, защищенность от волн и т.п., но все же невозможно представить, что каждая ассоциация встречается в особых условиях среды, в которых не могут доминировать другие виды. На мой взгляд, фитоценотическая замещаемость среди водных растений весьма высока.
Но почему такие фитоценотически замещающие виды, имеющие очень близкую экологию, не образуют смешанные, полидоминантные сообщества, в которых каждый из них имел бы примерно равную долю участия? Я думаю, причина существования монодоминантных сообществ водных растений заключается в том, что у многих видов водных растений преобладает вегетативное размножение. В результате чего образуются достаточно большие по площади и плотные клоны, которые геоботаники описывают как отдельные растительные сообщества. Какой вид будет доминировать в каждом конкретном месте, определяется случайностями в распространении вегетативных зачатков этих видов при заселении голого субстрата или при сукцессиях, связанных с изменениями среды. Победителем оказывается тот вид, который появляется там первым.
Вегетативное размножение растений и образование клонов играет важную роль в жизни многих типов растительных сообществ. На это обстоятельство геоботаники до последнего времени обращали мало внимания. Приведу лишь один пример. На западе Ленинградской обл., в Котельском заказнике есть обширное плоское плато, сложенное супесчаными почвами. На этом плато, на месте вырубленных около 40 лет назад лесов растут березовые леса, в травяном ярусе которых доминирует Calamagrostis arundinacea.
На однородном фоне вейниковых березняков нередко встречаются пятна диаметром несколько десятков метров, в которых преобладает Pteridium aquilinum. Такие пятна мы описывали как особые растительные сообщества. Когда позднее мы стали проводить классификацию березовых лесов, то оказалось, что березняки с доминированием орляка и вейника имеют полностью идентичный видовой состав. Их следует рассматривать как фитоценотически замещающие виды и объединять в одну ассоциацию. Известно, что орляк может образовывать очень крупные клоны, длина его корневищ может достигать 100 м и более.
Может ли такая ситуация иметь место в водных растительных сообществах? Я думаю, это вполне возможно. Тем более что площади многих водных сообществ невелики. В работе по водной растительности Западной Украины, выполненной недавно польскими геоботаниками (Swis, Soroka, 2000), приведены площади фитоценозов каждой ассоциации. Они, как правило, не превышают 10 м2. Такие небольшие по площади сообщества могут представлять собой клоны, возникшие в результате вегетативного размножения, но, разумеется, это не обязательно.
В монографии В. Г. Папченкова (2000) говорится о ненаправленных сменах, которые происходят на то обсыхающих, то обводняющихся мелководьях водохранилищ с переменным уровнем затопления. При таких сменах одно гидрофитное одновидовое или монодоминантное сообщество замещается на другое. Эти смены обусловлены потерей конкурентоспособности замещаемых популяций в связи с их возрастным состоянием, с цикличностью генеративного и вегетативного размножения, с поражением насекомыми, выеданием рыбами, птицами, млекопитающими и т.д. Эти смены могут быть обратимыми. Такие смены могут происходить только в том случае, когда образующие эти сообщества виды являются фитоценотически замещающими, когда они имеют сходную экологию.
Итак, и флористическая, и доминантная (эколого-фитоценотическая) классификации растительности имеют свои достоинства и недостатки. В условиях непрерывного и многомерного варьирования растительности ни одна из них не может дать дискретных и естественных синтаксонов. При классификации одновидовых или монодоминантных сообществ различия между этими классификациями стираются, так как нет другой основы, кроме доминантов, для выделения синтаксонов. Выделение более высоких единиц классификации производится на основе сходства жизненных форм доминирующих видов, и так строится классификация водной растительности независимо от того, принципов какой школы придерживаются исследователи.
Следует остановиться еще на одной проблеме, которая возникает при построении классификации, охватывающей обширные территории. Речь идет о географически замещающих видах. Приведу лишь два примера. С ними мы столкнулись при проведении классификации еловых лесов европейской России. Формально на этой территории ельники образованы двумя видами ели: Picea abies и P. obovata. На большей части территории Европейской России преобладают формы, переходные между этими двумя видами.
Кроме того, нет никаких убедительных свидетельств, что эти виды чем-то отличаются по своей экологии, обладают каким-то отличным средообразующим воздействием. В этом случае правильнее рассматривать все леса с доминированием этих двух видов ели как одну формацию или союз в рамках флористической классификации, как в свое время сделал J. Braun-Blanquet. Сравнивая леса с этими двумя видами ели, мы не нашли никаких заметных различий между ельниками на западе и востоке Европейской России.
На востоке Европейской России Calamagrostis arundinacea постепенно замещается С. obtusata. Эти два вида также имеют сходную экологию, и вряд ли стоит выделять разные ассоциации на основе доминирования того или иного вида. Эти примеры не могут служить основой для общего правила. В каждом случае вопрос с учетом при классификации географически замещающих видов должен решаться отдельно.
А как быть с теми мелкими видами, которых становится все больше в наших «Флорах»? К сожалению, и здесь однозначного ответа быть не может. Вряд ли геоботаники будут использовать виды Taraxacum, выделенные из T. officinale, или из группы Ranunculus auricomus. Но нельзя рассматривать при классификации растительности все виды манжеток как один вид Alchemilla vulgaris, так как среди них имеются виды с очень разной экологией. В то же время нередко в западноевропейских работах в качестве дифференциальных видов используются подвиды, но они имеют различия или в экологии, или в географических ареалах.
Очень серьезный недостаток многих работ по классификации растительности, независимо от используемых принципов, состоит в том, что выделяемые синтаксоны недостаточно глубоко обосновываются. Недостаточно перечислить доминирующие виды в каждом ярусе или указать 2—3 дифференциальных видов. Нередко при проведении доминантной классификации возводят в ранг особых синтаксонов группы сообществ, отличающиеся только соотношением обилий видов.
Такие группы могут определяться лишь случайным варьированием. Что очень часто отсутствует в работах по классификации, так это серьезное обоснование выделенных единиц. И этот недостаток свойствен работам с использованием любых принципов классификации растительности. Формальное проведение классификации, увлечение вопросами терминологии, отсутствие сравнения единиц друг с другом, с материалами, опубликованными по другим районам, — именно это делает классификацию неудовлетворительной.
Построению обоснованной классификации растительности мешает недостаточная изученность экологии и биологических особенностей многих видов растений. Специальных работ в этом направлении ведется довольно мало, но только такие работы помогут оценить специфику средообразующей роли вида, его индикаторное значение, место в сукцессионных сменах. Работа выполнена при поддержке Российского фонда фундаментальных исследований (грант 01-04-49629).
Дата добавления: 2023-05-30; просмотров: 985;
Поиск по сайту
Узнать еще
- Анемии, связанные с нарушением включения железа в синтез гема и с дефектами утилизации порфиринов
- Археология или история материальной культуры?
- Атомные модели дефектов. Нульмерные, или точечные дефекты
- Заготовки кормов. Сушка сена активным вентилированием
- Классификация неоплазм и онкологическая терминология
- Классификация рефлекторных реакций. Порядок изучения прирожденной и индивидуально приобретенной деятельности
- Классификация соединений костей скелета
- Место, занимаемое печью или камином. Кладка корпуса

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
