Вопрос 2. Химизм дыхания
Дыхание состоит из двух этапов:
· анаэробный (гликолиз), в результате которого углеводы распадаются до пировиноградной кислоты (пируват, ПВК).
· аэробный – окисление ПВК до СО2 и Н2О.
При недостатке кислорода (в анаэробных условиях) ПВК расщепляется не до СО2 и Н2О, а до спирта или других соединений в процессе брожения (см. раздел «Микробиология»)
Анаэробная фаза (гликолиз) общий начальный этап аэробного типа дыхания и всех видов брожений, представлен во всех формах жизни, и предшествует всем типам дыхания, протекает в цитоплазме. В результате гликолиза молекула глюкозы расщепляется на две молекулы пирувата (ПВК).
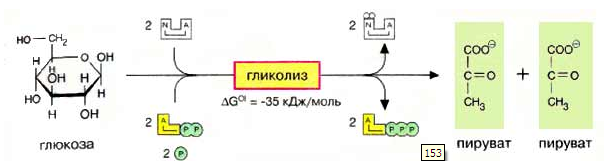
При расщеплении образуется 2 молекулы АТФ и восстанавливается 2 молекулы НАДФ·Н2. При окислении молекула НАДФ·Н2 даёт 3 молекулы АТФ. Ззапас энергии в макроэргических связях АТФ равен примерно 40 кДж. Таким образом, суммарный выход энергии гликолиза равен 320 кДж.
Значение гликолиза для растения: является источником энергии; в процессе гликолиза образуются промежуточные продукты окисления, необходимые для биосинтеза других сложных органических соединений (амино- и жирных кислот, фенольных соединений, лигнина).
Аэробная фаза осуществляется с участием комплекса ферментов, содержащихся в митохондрии. ПВК, образуемая при гликолизе, в цитоплазме связывается с коферментом А с образованием ацетилкофермента А (ацетил-КоА), который поступает в митохондрии и вступает в цикл Кребса. Ферменты цикла расположены в белковом матриксе. На внутренней мембране локализованы ферменты ЭТЦ (электронно-транспортная цепь). В результате последовательных превращений в аэробных условиях ПВК окисляется до конечных, самых простых, предельно окисленных продуктов – СО2 и Н2О.
Цикл Г.А. Кребса (цикл трикарбоновых кислот – ЦТК) протекает без кислорода воздуха, но с участием кислорода воды(мокрое окисление) встроме митохондрий. При этом ПВК взаимодействует со ЩУК(щавелевоуксусная кислота) и включается в цепь последовательных превращении (реакции отнятия и присоединения воды, декарбоксилирования и дегидрирования). с образованием ди- и трикарбоновых кислот: лимонная→цис-аканитовая→изолимонная→ЩЯК (щавелевоянтарная кислота)→ά-кето-глутаровая→янтарная→фумаровая→яблочная→ЩУК (см. рисунок). Суммарное уравнение цикла Кребса: 2СН3-СО-СООН + 5О → 6СО2 + 4Н2О
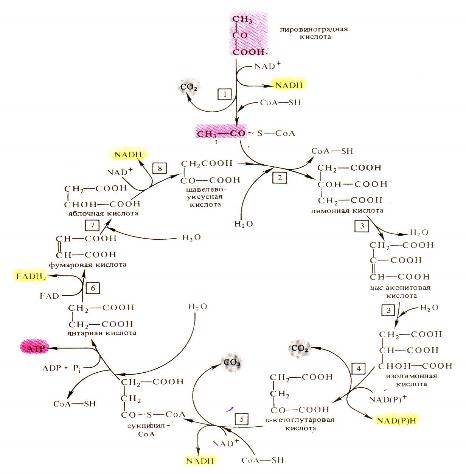
| Рисунок – Аэробная фаза дыхания (цикл Кребса) |
Заключительным этапом дыхания является окислительное фосфорилирование восстановленных в цикле Кребса коферментов. В результате которого происходит отнятие электронов от NADH, NAD(P)H и FADН2 и последовательная передача их через цепь переносчиков по ЭТЦ на кислород, в результате чего высвобождается энергия. Переносчики ЭТЦ расположены в порядке убывания их окислительно-восстановительного потенциала (от -0,32 у дыхательного субстрата до +0,82 у О2). Электроны «скатываются» с «энергетической» горки благодаря разности потенциалов не сразу, а постепенно, что позволяет, избежать неэффективного выброса энергии и связать её в форме АТФ. Движение электронов по ЭТЦ вызывает перекачивание протонов через внутреннюю мембрану из матрикса митохондрии в межмембранное пространство, это приводит к возникновению градиента концентрации протонов (в матриксе рН ниже) и созданию разности потенциалов на мембране (внутренняя сторона мембраны заряжается «–», а наружная – «+»). Обе силы стремятся переместить протоны обратно в матрикс митохондрии, но внутренняя мембрана для них непроницаема. Обратный транспорт протонов возможен только через особые белковые комплексы на мембране – АТФ-синтетазы. Когда протоны проходят по трансмембранному каналу АТФ-синтетазы из межмембранного пространства в матрикс митохондрии, то при транспорте каждых 2-х протонов синтезируется 1 молекула АТФ. Таким образом, синтез АТФ сопряжен с транспортом электронов и перекачиванием протонов.
Энергетический выход дыхания. В цикле Кребса из 1 молекулы ПВК при окислении образуется 3 молекулы СО2, 1 молекула NAD(P)H, 3 молекулы NADH, 1 молекула FADН2 и 1 молекула АТФ синтезируется в ходе субстратного фосфорилирования. В энергетическом отношении 1 молекула NAD(P)H или NADH соответствует 3 молекулам АТФ, а молекула FADН2 – 2 молекулам АТФ. Тогда в итоге образуется 15 молекул АТФ, а поскольку в цикле участвуют 2 молекулы ПВК – то всего синтезируется 30 молекул АТФ (1200кДж). При гликолизе было образовано 8 молекул АТФ (320 кДж). Тогда суммарная энергетика процесса дыхания составляет 1520 кДж в виде АТФ. Всего при дыхании образуется 2874 кДж энергии, из которых 1354 кДж распределяется в виде тепла. Таким образом, КПД дыхания составляет 54%. Это очень много, такого КПД нет ни в одном двигателе внутреннего сгорания.
В аэробных условиях основным источником энергии в клетке служит окислительное расщепление дыхательных субстратов, а в отсутствие кислорода – гликолиз и брожение. Поэтому на уплотнённых, переувлажнённых почвах продуктивность растений снижается, так как наблюдается истощение растений (энергетический эффект брожения в 25 раз ниже дыхания) и отравление продуктами брожения (СО2, кислоты, спирты).
Вопрос 3. Зависимость дыхания от внутренних и внешних факторов
Биологические особенности. Очень низкая интенсивность дыхания у суккулентов, несколько выше – у хвойных, а самая высокая – у плесневых грибов. Интенсивнее дышат светолюбивые и быстрорастущие растения. В онтогенезе растения интенсивность дыхания сначала нарастает, достигает мах (период зрелости), а затем снижается. Сухие семена дышат слабо, однако по мере их набухания интенсивность дыхания увеличивается, достигая наибольших значений в период интенсивного роста зародыша. Дыхание листьев и корней, максимально в молодом возрасте, снижается по мере их старения. На растении интенсивнее дышат репродуктивные органы, нежели вегетативные, в плодах и семенах кожура дышит интенсивнее мякоти. Интенсивность дыхания также повышается при повреждении или инфицировании растения.
Содержание углеводов. Углеводы – основной субстрат дыхания, поэтому интенсивность дыхания тесно коррелирует с их содержанием, особенно в молодых растущих органах. После завершения роста и в стареющих тканях, корреляция между указанными параметрами быстро ослабевает.
Содержание воды.С усилением дефицита влаги подавляется, прежде всего, рост, затем фотосинтез и в последнюю очередь дыхание. При быстром падении влагообеспеченности, интенсивность дыхания сначала может даже возрастать (увеличивается коэффициент дыхания на поддержание при относительном постоянстве коэффициента на рост). При постепенном нарастании дефицита влаги, дыхание уменьшается параллельно снижению скорости роста и фотосинтеза. Наиболее чувствительным к усилению дефицита влаги в почве является дыхание стеблей, затем листьев и колосьев пшеницы. ИД зерна существенно зависит от его влажности, имея минимальные значения при критической влажности. Это необходимо учитывать при хранении зерна.
Важно также содержание воды в самом растении. Сухие семена при повышении влажности дышат интенсивнее, а при созревании семян дыхание затухает. У сочной растительности и плодов увядание сопровождается усилением дыхания. Эти особенности следует учитывать при хранении продукции: зерно следует хранить при стандартной влажности – 12-14%, у влажного зерна за счет интенсивного дыхания повышается температура – зерно «сгорает», теряет всхожесть; сочную продукцию хранят при повышенной влажности (80-95%), не допуская увядания.
Температура.Влияние температуры на интенсивность дыхания подчиняется правилу Вант-Гоффа: с повышением температуры дыхательная активность возрастает (при увеличении температуры на 100С интенсивность дыхания удваивается). При очень высоких температурах наблюдается нарушение обмена веществ и дыхание быстро падает. Координальные точки дыхания: tmin=0, tмах=600С, topt=40 (если ткань долго выдерживать при такой t, то topt падает до 25). В диапазоне 0-400 интенсивность дыхания возрастает, 40-600 – замедляется, а при t > tмах – прекращается.В условиях постоянной температуры дыхание ниже, чем при переменных. Таким образом, хранить продукцию следует при невысокой постоянной температуре.
Концентрация СО2 и О2.При достаточном содержании О2 дыхание возрастает. Однако, оно заметно снижается в области как очень высоких (100%), так и низких (3% и ниже) концентраций О2. Последнее наблюдается при избыточном переувлажнении и уплотнении почвы, в зимний период при выпревании и образовании ледяной корки. Растения истощаются и погибают от отравления продуктами брожения. Обогащение атмосферы СО2 снижает интенсивность дыхания, что используют при длительном хранении плодов и овощей в регулируемой газовой среде.
Свет. Интенсивность дыхания на свету выше, чем в темноте. При этом усиление дыхания вызывает синий свет, поглощаемый флавинами – компонентами дыхательной цепи, красный свет влияния не оказывает. Свет оказывает также и косвенное влияние на дыхание. При длительном нахождении растения в темноте наблюдается недостаток дыхательного субстрата, который образуется в процессе фотосинтеза на свету.
Минеральное питание.Внесение минеральных веществ в почву, как правило, сопровождается усилением дыхания корней. Это связано с повышением затрат энергии на транспорт ионов. Наиболее интенсивно дыхание корней возрастает при внесении азотных удобрений, так как кроме транспорта ионов, энергия затрачивается на восстановление нитратов. Значительно выше энергетические затраты на связывание атмосферного азота клубеньками бобовых, дыхание клубеньков во много раз выше, чем корней. Интенсивность дыхания корней с клубеньками у бобовых культур при недостатке N в 2 раза выше, чем на высоком фоне.
ТЕМА 7
Дата добавления: 2016-12-16; просмотров: 6643;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
