Скелет пресмыкающихся: сравнительная анатомия и особенности строения
Систематическое положение объектов изучения включает несколько отрядов класса Пресмыкающиеся (Reptilia). В качестве основных представителей рассматриваются: серый варан (Varanus griseus Daud.) из отряда Чешуйчатые (Squamata), миссисипский аллигатор (Alligator mississippiensis Daud.) из отряда Крокодилы (Crocodilia) и болотная черепаха (Emys orbicularis L.) из отряда Черепахи (Chelonia или Testudines). Изучение скелетов этих животных позволяет проследить основные направления эволюционных изменений в пределах класса и выявить адаптации к различным средам обитания.
Материал и оборудование для практического занятия включают смонтированные скелеты варана, змеи и черепахи, а также черепа крокодилов (или их муляжи). Для детального анализа на одного-двух студентов предусмотрен разборный скелет варана, череп черепахи и препаровальные иглы. В ходе работы необходимо рассмотреть общее строение скелета варана, его основные отделы, а также особенности скелета змеи и черепахи, сравнить строение черепов различных представителей и выполнить схематические рисунки ключевых структур.
Осевой скелет (позвоночник). Общая характеристика позвоночника пресмыкающихся отличается более четкой дифференцировкой на отделы по сравнению с земноводными. Выделяют шейный, грудопоясничный, крестцовый и хвостовой отделы. Позвонки у большинства рептилий процельные (тело позвонка вогнуто спереди и выпукло сзади), что обеспечивает прочное и гибкое соединение. Над телом позвонка располагаются верхние дуги, образующие позвоночный канал, и остистый отросток, а также передние и задние сочленовные отростки, которые увеличивают прочность позвоночника при изгибах.
Шейный отдел (pars cervicalis) состоит из нескольких позвонков, причем первые два имеют уникальное строение. Первый шейный позвонок — атлант (atlas) — лишен тела и имеет форму кольца, разделенного на две части. Второй позвонок — эпистрофей (epistropheus) — несет спереди зубовидный отросток (processus odontoideus), представляющий собой тело атланта, сросшееся с эпистрофеем. Этот отросток свободно входит в отверстие атланта, обеспечивая вращение головы. Остальные шейные позвонки несут короткие шейные ребра.
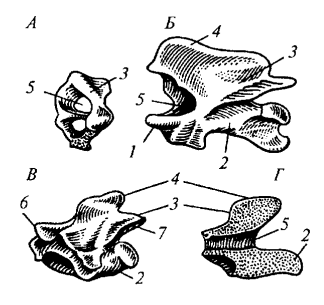
Рис. 81. Позвонки варана: А — атлант; Б — эпистрофей; В — грудной позвонок; Г— продольный разрез грудного позвонка: 1 — зубовидный отросток эпистрофея; 2 — тело позвонка; 3— верхняя дуга; 4— остистый отросток; 5 — канал для спинного мозга; 6 — передний сочленовный отросток; 7— задний сочленовный отросток
Грудной и поясничный отделы часто объединяют в грудопоясничный (pars thoracolumbalis). В грудном отделе ребра соединяются с грудиной, формируя грудную клетку; в поясничном отделе ребра короткие и не доходят до грудины. К позвонкам причленяются ребра, которые у амниот становятся двухзвенными. Крестцовый отдел (pars sacralis) включает два позвонка с мощными поперечными отростками, к которым прикрепляются кости таза. Хвостовой отдел (pars caudalis) состоит из многочисленных, постепенно уменьшающихся позвонков.
Особенности строения позвоночника у змей связаны с редукцией парных конечностей и специфическим способом передвижения. Позвоночник отчетливо делится только на туловищный и хвостовой отделы. Все туловищные позвонки несут длинные подвижные ребра, которые свободны (грудина отсутствует) и упираются в брюшные роговые щитки, обеспечивая движение.
Особенности строения позвоночника у черепах определяются их уникальным панцирем. Туловищные позвонки, ребра и кожные кости срастаются, образуя карапакс (спинной щит). Тела позвонков и ребра расширены и уплощены, они неподвижно соединены с костными пластинками покровного происхождения. Шейный и хвостовой отделы остаются подвижными, причем шейные позвонки имеют смешанный тип строения: передние — опистоцельные (выпуклые спереди и вогнутые сзади), задние — процельные, а между ними находится позвонок с двояковыпуклым телом.
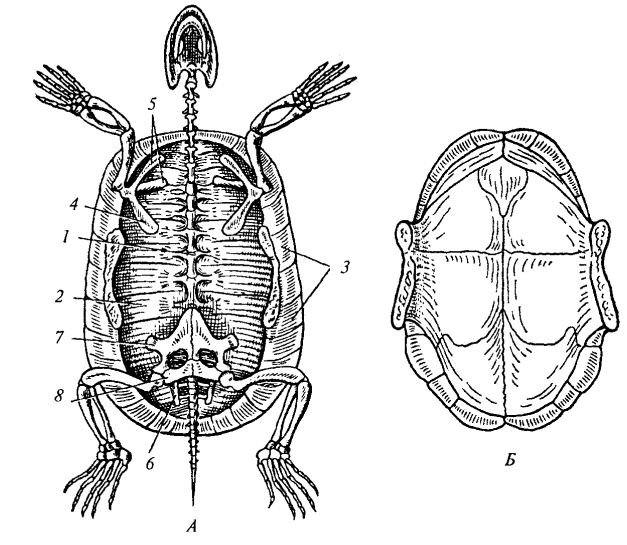
Рис. 82. Скелет болотной черепахи: А — карапакс; Б — пластрон: 1 — туловищный отдел позвоночного столба; 2 — реберные пластинки; 3— краевые пластинки; 4 — коракоид; 5 — лопатка; 6 — подвздошная кость; 7 — лобковая кость; 8 — седалищная кость
Череп пресмыкающихся. Общая характеристика черепа рептилий отличается более полным окостенением по сравнению с амфибиями. Хрящ сохраняется лишь в обонятельной капсуле и слуховой области. Череп состоит из осевого (мозгового) и висцерального отделов, которые у взрослых животных образуют единое целое. В его состав входят как замещающие (хрящевые), так и покровные (кожные) кости. В качестве модели для изучения удобно использовать череп варана.
Осевой череп варана включает затылочный отдел из четырех костей: основной затылочной (basioccipitale), двух боковых затылочных (occipitale laterale) и верхней затылочной (supraoccipitale). Они окружают большое затылочное отверстие и образуют единственный затылочный мыщелок (condylus occipitalis), сочленяющийся с атлантом. В слуховом отделе самостоятельна только переднеушная кость (prooticum). В основании черепа лежит основная клиновидная кость (basisphenoideum), а спереди — парные сошники (vomer). Крышу черепа формируют покровные кости: теменные (parietale), лобные (frontale), носовые (nasale), предлобные (prefrontale), предглазничные (preorbitale) и слезные (lacrimale).
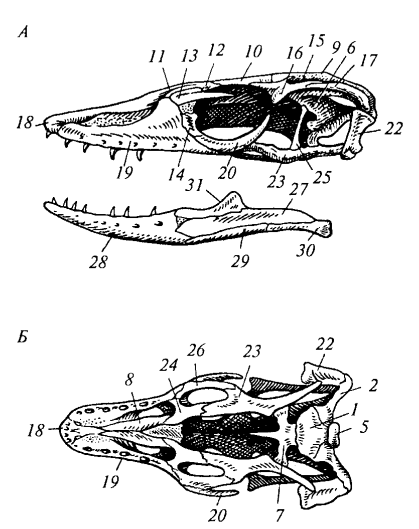
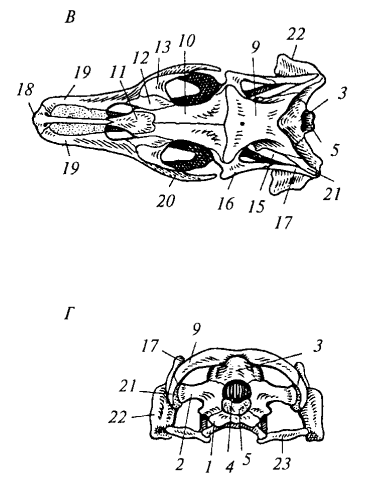
Рис. 83. Череп варана: А — сбоку; Б — снизу; В — сверху; Г — сзади: 1 — основная затылочная кость; 2 — боковая затылочная кость; 3 — верхняя затылочная кость; 4 — большое затылочное отверстие; 5— затылочный мыщелок; б — переднеушная кость; 7 — основная клиновидная кость; 8 — сошник; 9 — теменная кость; 10 — лобная кость; 11 — носовая кость; 12 — предлобная кость; 13 — пред- глазничная кость; 14 — слезная кость; 15 — верхняя височная яма; 16 — заглазничная кость; 17 — чешуйчатая кость; 18— предчелюстная кость; 19 — верхнечелюстная кость; 20 — скуловая кость; 21 — надвисочная кость; 22— квадратная кость; 23 — крыловидная кость; 24— нёбная кость; 25 — верхнекрыловидная кость; 26— поперечная кость; 27 — надугловая кость; 28 — зубная кость; 29 — угловая кость; 30— сочленовная кость; 31 — венечная кость
Височные дуги и типы черепа. У рептилий в связи с редукцией сплошного костного панциря (стегального черепа) формируются височные ямы и ограничивающие их височные дуги. По количеству дуг различают несколько типов черепа. Диапсидный (двудужный) череп характерен для крокодилов, ящериц и змей, но степень развития дуг варьирует. У крокодила (рис. 84) имеются две полные височные дуги: верхняя (из заглазничной и чешуйчатой костей) и нижняя (из скуловой и квадратно-скуловой костей). У варана нижняя дуга редуцирована (отсутствует квадратно-скуловая кость), поэтому боковая височная яма остается открытой. У змей (рис. 86) редуцированы обе дуги, что обеспечивает чрезвычайную подвижность челюстного аппарата.
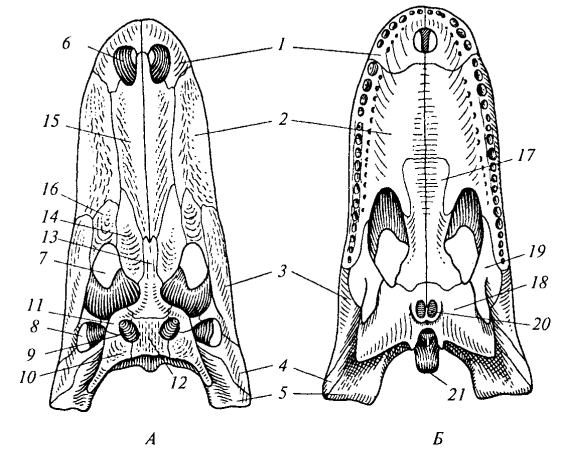
Рис. 84. Череп крокодила (миссисипский аллигатор): А — сверху; Б — снизу: 1 — предчелюстная кость; 2 — верхнечелюстная кость; 3 — скуловая кость; 4 — квадратно-скуловая кость; 5 — квадратная кость; 6 — наружная ноздря; 7 — глазница; 8 — боковая височная яма; 9 — верхняя височная яма; 10 — чешуйчатая кость; 11 — заглазничная кость; 12 — теменная кость; 13 — лобная кость; 14 — предлобная кость; 15 — носовая кость; 16 — слезная кость; 17— нёбная кость; 18 — крыловидная кость; 19 — поперечная кость; 20— хоаны; 21 — затылочный мыщелок
У черепах (рис. 85) височные ямы отсутствуют, а боковая стенка образована плотно сросшимися костями, формирующими ложную височную яму. Такой тип черепа называется анапсидным (бездужным). Он считается примитивным и унаследован от древних рептилий.
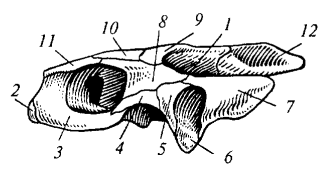
Рис. 85. Череп болотной черепахи: 1 — ложная височная яма; 2 — предчелюстная кость; 3 — верхнечелюстная кость; 4 — скуловая кость; 5 — квадратно-скуловая кость; 6 — квадратная кость; 7 — чешуйчатая кость; 8 — заглазничная кость; 9 — теменная кость; 10 — лобная кость; 11 — предлобная кость; 12 — верхняя затылочная кость
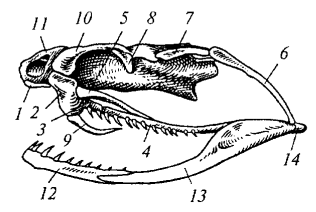
Рис. 86. Череп ядовитой змеи: 1 — предчелюстная кость; 2 — верхнечелюстная кость; 3 — нёбная кость; 4 — крыловидная кость; 5 — поперечная кость; б — квадратная кость; 7 — над- височная кость; 8— заднелобная кость; 9— ядовитый зуб; 10 — предлобная кость; 11 — носовая кость; 12— зубная кость; 13 — угловая кость; 14 — сочленовная кость
Висцеральный череп включает производные жаберных дуг. У варана нёбноквадратный хрящ окостеневает, образуя квадратную кость (quadratum), к которой подвижно причленяется нижняя челюсть. Впереди лежат крыловидная (pterygoideum), нёбная (palatinum) кости, а также верхнекрыловидная (epipterygoideum) и поперечная (transversum) кости. Вторичная верхняя челюсть представлена предчелюстными (premaxillare) и верхнечелюстными (maxillare) костями. Нижняя челюсть состоит из замещающей сочленовной кости (articulare) и покровных: зубной (dentale), угловой (angulare), надугловой (supraangulare), венечной (coronare) и других.
Зубы у большинства рептилий (кроме черепах) простые конические, прирастающие к краю челюстей. У крокодилов зубы текодонтные — сидят в отдельных ячейках (альвеолах). У змей имеются специализированные ядовитые зубы с каналом или бороздкой для стекания яда. Кроме того, у змей чрезвычайно подвижны квадратные, крыловидные, нёбные и верхнечелюстные кости, что позволяет заглатывать крупную добычу. У крокодилов развито вторичное твердое нёбо, отделяющее носоглоточный ход от ротовой полости и позволяющее дышать при заполненном водой рте.
Парные конечности и их пояса. Плечевой пояс пресмыкающихся состоит из лопатки (scapula) и коракоида (coracoideum), которые участвуют в образовании суставной впадины для передней конечности. Лопатка продолжается надлопаточным хрящом, а спереди к коракоиду примыкает прокоракоидный хрящ. Грудина (sternum) хорошо развита, к ней причленяются грудные ребра, формирующие грудную клетку. На вентральной стороне грудины расположен Т-образный надгрудинник (episternum) — покровная кость, а впереди него — ключицы (clavicula), соединяющиеся с лопатками и надгрудинником.
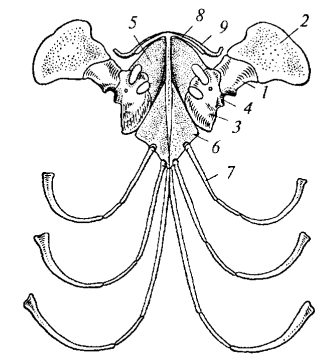
Рис. 87. Плечевой пояс варана (вид снизу): 1 — лопатка; 2 — надлопаточный хрящ; 3 — коракоид; 4 — суставная впадина для головки плеча; 5 — про- коракоидный хрящ; 6 — грудина; 7 — ребра; 8 — надгрудинник; 9 — ключица
У змей плечевой пояс полностью редуцирован. У черепах ключицы и надгрудинник входят в состав пластрона (брюшного щита панциря), образуя передние парные и непарную костные пластинки.
Тазовый пояс состоит из двух симметричных половин, каждая из которых включает три кости: подвздошную (ilium), лобковую (pubis) и седалищную (ischium). Все они участвуют в образовании вертлужной впадины (acetabulum) для задней конечности. Таз у рептилий закрытый: правая и левая лобковые и седалищные кости срастаются на брюшной стороне.
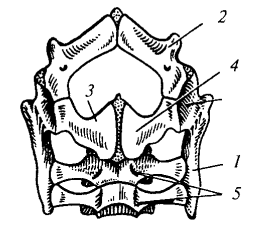
Рис. 88. Тазовый пояс варана (вид снизу): 1 — подвздошная кость; 2 — лобковая кость; 3 — седалищная кость; 4 — вертлужная впадина (сочленовная ямка) для головки бедра; 5— крестцовые позвонки
Строение передней конечности (рис. 89, А) включает плечевую кость (humerus), предплечье из локтевой (ulna) и лучевой (radius) костей, и кисть. Кисть состоит из запястья (carpus), пясти (metacarpus) и фаланг пальцев. Характерной особенностью является интеркарпальный сустав, проходящий между проксимальным и дистальным рядами костей запястья, что увеличивает подвижность кисти.
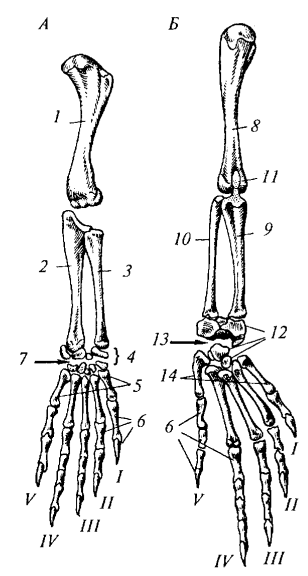
Рис. 89. Конечности варана: А — передняя; Б — задняя: 1 — плечевая кость; 2 — локтевая кость; 3 — лучевая кость; 4 — запястье; 5 — пясть; 6 — фаланга пальцев; 7— интеркар- пальный сустав; 8 — бедренная кость; 9 — большая берцовая кость; 10 — малая берцовая кость; 11— коленная чашечка; 12 — предплюсна; 13 — интертарзальный сустав; 14 — плюсна
Строение задней конечности (рис. 89, Б) включает бедренную кость (femur), голень из большой берцовой (tibia) и малой берцовой (fibula) костей, и стопу. В коленном суставе имеется коленная чашечка (patella). В стопе выделяют предплюсну (tarsus), плюсну (metatarsus) и фаланги пальцев. Для рептилий характерен интертарзальный сустав между проксимальным и дистальным рядами костей предплюсны. Все пальцы заканчиваются когтями.
Заключение: прогрессивные черты организации рептилий. Адаптации к наземному образу жизни. Рептилии — первый класс настоящих наземных позвоночных (Amniota). Их организация характеризуется комплексом приспособлений, направленных на экономию влаги и независимость от водной среды. Кожа сухая, лишенная желез, покрыта роговыми чешуями, предохраняющими от высыхания. Легкие имеют сложное губчатое строение с большой дыхательной поверхностью, а вентиляция осуществляется за счет движения грудной клетки (реберное дыхание).
Прогресс выделительной и кровеносной систем. У рептилий функционируют тазовые почки (metanephros), канальцы которых полностью утратили связь с полостью тела. Обратное всасывание воды в почечных канальцах позволяет экономить влагу. В кровеносной системе появляется неполная перегородка в желудочке, что обеспечивает частичное разделение потоков артериальной и венозной крови. Голова и передние конечности снабжаются почти чистой артериальной кровью. У крокодилов желудочек разделен полностью (четырехкамерное сердце), но сохраняются обе дуги аорты с перекрестом.
Эволюция скелета и нервной системы. В скелете важнейшим приобретением является формирование грудной клетки, обеспечивающей не только эффективное дыхание, но и прочную опору для плечевого пояса. Подвижность головы увеличилась благодаря единственному затылочному мыщелку и специализации первых двух шейных позвонков. В черепе происходит редукция сплошного костного панциря и образование височных ям, что облегчает череп и создает пространство для челюстной мускулатуры. Головной мозг относительно крупнее, чем у амфибий; полушария переднего мозга имеют кору и ассоциативные центры.
Разнообразие жизненных форм. Рептилии освоили разнообразные среды обитания: пустыни, леса, скалы, пресные водоемы и даже море. Среди них есть хищники, растительноядные (наземные черепахи) и формы со специализированными приспособлениями (ядовитые зубы, покровительственная окраска, автотомия хвоста). Вторичноводные виды (крокодилы, морские черепахи, змеи) сохраняют связь с сушей для размножения, а некоторые (морские змеи) перешли к яйцеживорождению.
Эволюционное значение. Расцвет рептилий пришелся на мезозойскую эру, когда они занимали господствующее положение. Однако пойкилотермность (неспособность поддерживать постоянную температуру тела) ограничила их распространение в эпоху похолодания, что привело к вытеснению более прогрессивными группами — птицами и млекопитающими. Тем не менее, современные рептилии сохраняют важное значение в экосистемах и представляют огромный интерес для изучения эволюционных процессов.
Рекомендуемая литература: 1. Банников А. Г., Даревский И. С., Ищенко В. Г. и др. Определитель земноводных и пресмыкающихся СССР. М., 1977.
2. Гуртовой Н. Н. Систематика и анатомия хордовых животных: Краткий курс. М., 2004.
3. Гуртовой Н. Н., Матвеев Б. С., Дзержинский Ф. Я. Практическая зоотомия позвоночных. Земноводные, пресмыкающиеся. М., 1978.
4. Дзержинский Ф. Я. Сравнительная анатомия позвоночных животных. М., 1998.
5. Ромер А., Парсонс Г. Анатомия позвоночных: В 2 т. М., 1992.
Сведения об авторах и источниках:
Авторы: Карташев Н. Н., Соколов В. Е., Шилов И. А.
Источник: Практикум по зоологии позвоночных
Данные публикации будут полезны студентам биологических специальностей, изучающих зоологию и сравнительную анатомию, а также всем, кто интересуется эволюционной морфологией и строением примитивных хордовых животных.
Дата добавления: 2026-03-05; просмотров: 116;
Поиск по сайту
Узнать еще
- Активные и пассивные операции Сберегательного банка России: структура и особенности
- Анатомические и физиологические особенности птиц. Перьевой покров
- Анатомия акулы: подробное руководство по вскрытию и систематике
- Анатомия земноводных: методика вскрытия и особенности внутреннего строения на примере травяной лягушки (Rana temporaria L.)
- Анатомия и физиология пищеварительной и мочеполовой системы бесхвостых амфибий на примере лягушки
- Анатомия костистых рыб: методика вскрытия и особенности внутреннего строения на примере щуки (Esox lucius L.)
- Анатомия млекопитающих: полное руководство по вскрытию белой крысы
- Анатомия пресмыкающихся: вскрытие кавказской агамы (Agama caucasica) и особенности строения рептилий

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
