Тема: “Промысловая смертность рыб”.
Вылов рыбы и других морепродуктов неуклонно растет. Если в начале века он составлял 10 млн. т, то сейчас превышает 100 млн. т . Одной из причин роста мировой добычи рыбы можно считать повышение уловистости орудий и эффективности способов лова. Уловистостью называется способность орудий лова улавливать рыбу или нерыбные объекты. Уловистость зависит от орудия лова, объекта лова, условий.
Под уловистостью, или абсолютной уловистостью, понимается отношение количества добытой рыбы к количеству, находившемуся в зоне облова отцеживающим орудием, или отношение количества рыбы, попавшей в сети либо ставную ловушку, к количеству рыбы, прошедшей через сечение, занятое орудием лова.
Под относительной уловистостью понимают отношение количества рыбы, добытой данным орудием, к количеству рыбы, добытой за тот же срок в аналогичных сходным по величине орудием, но иной конструкцией или сделанным из иных материалов.
Уловистость орудий лова и эффективность способа лова зависят как от конструкции орудия и техники лова, так и в не меньшей степени от приспособительных свойств облавливаемой популяции рыб. Уловистость орудия лова – это результат взаимодействия его, популяции облавливаемой рыбы и условий лова.
Уловистость орудия лова будет зависеть от следующих факторов:
1. конструкция, материал орудия и способ лова;
2. незаметность орудия лова для рыбы;
3. характер поведения рыбы в зоне облова;
4. гидрометеорологическая обстановка в районе лова.
Существенное влияние оказывает правильное использование орудия лова. Например, при лове ставными сетями, при правильной расстановке сетей можно значительно повысить уловы.
Уловистость крючковых наживных орудий зависит не только от конструкции поводцев, но и от качества наживки или ее имитации, суточного ритма питания рыбы.
Для сравнения уловистости различных орудий лова вводят коэффициенты уловистости. Для закидных неводов они колеблются от 0,37 до 0,67. У тралов, кошельковых неводов другие значения.
Уловистость крючковых орудий лова сильно зависит от накормленности рыбы.
Уловистость может зависеть от возрастного состава облавливаемого стада. В высокоурожайные годы уловистость увеличивается и наоборот.
Реакция рыбы на приближение орудия лова имеет общие черты с ее реакцией на хищника. Поэтому важна скорость и незаметность орудия лова. Для характеристики уловистости иногда пользуются другими понятиями – селективность, удерживающая способность орудия.
Селективность орудий и способов лова. Под селективностью понимается способность орудия лова улавливать рыбу с определенной характеристикой, поэтому, ее иногда называют избирательной способностью.
Всякий вылов в той или иной степени и форме селективен. В некоторых случаях селективность рыболовства может быть выражена очень слабо, и практически ее можно не учитывать, в других (таких случаев большинство) селективность рыболовства сильно отражается на структуре стада. Селективность рыболовства определяется двумя моментами: отбирающей способностью орудий рыболовства и поведением рыб ( разного пола, разного физиологического состояния, разного возраста).
Характер отбирающего действия орудия рыболовства есть результат взаимодействия орудия и рыбы.
При изменении ячеи сетного полотна изменяется минимальный размер залавливаемых орудием рыб. На правую часть размерного ряда облавливаемой популяции рыб изменение размеров ячеи (до определенных пределов) не оказывает прямого влияния, и обычно правая ветвь кривой размерного состава рыб в уловах отражает размерный состав более крупных рыб в облавливаемой популяции. При лове объячеивающими орудиями существует прямая связь между минимальным, средним и максимальным размерами добываемых рыб и размерами ячеи сетного полотна.
Особенно детальные исследования проведены по изучению селективности кутцов тралов. А.И. Трещев (1964г.) показал, что связь размеров ячеи и длины рыбы выражается простой формулой:
а = k * l ,
где а – размер ячеи (шаг);
l – размер рыбы;
k – коэффициент, который зависит от вида и экстерьера рыбы и меняется с возрастом и изменением ее упитанности.
По Ф.И. Баранову:
a = k1 L ; a = k2 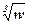
где k1 – линейный коэффициент;
k2 – массовый коэффициент;
w – масса рыбы.
Под коэффициентом селективности (ks) обычно понимается отношение длины рыбы (l), 50% которой проходит через ячею, а 50% остается в орудии лова, к размерам ячеи (В), т.е.:
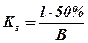
Интенсивность рыболовства и степень его воздействия на стадо промысловой рыбы зависит от очень многих причин, которые далеко не поддаются точному учету. Различают абсолютную и относительную интенсивность рыболовства.
В общей форме абсолютная интенсивность рыболовства – это произведение величины промыслового усилия на коэффициент уловистости и выражается формулой:
J = U * g ,
где U – промысловое усилие;
g - коэффициент уловистости.
Промысловое усилие – это улов за 100 часов работы орудия лова.
Относительная интенсивность рыболовства – это процент изымаемой части стада от всего стада, достигшего промысловых размеров.
Естественно, что успешность рыболовства определяется не только уловистостью орудий и эффективностью способов лова, но и тем, насколько районы облова совмещаются с местами концентрации объектов лова.
Место скоплений находят на основе знания поведения и распределения рыбы (на промысловых картах).
Существенную роль играет и непосредственное обнаружение рыбы по всплескам (тунцы), по скоплению птиц, непосредственно с самолета (стаи сардин, скумбрии, анчоусов и др.).
Разная интенсивность вылова самцов и самок может явиться результатом неодинаковой реакцией особей разных полов на один и тот же раздражитель. Так, самки кильки с текучими половыми продуктами не привлекаются светом, и ускользают от облова.
Объячеивающие орудия лова могут отбирать из популяции рыб определенного экстерьера, т.е. определенной стадии зрелости (с отвисшим брюшком).
На примере сельдевых, карповых, а затем окуневых показано, что сети отбирают из популяции наиболее упитанных, но и мелких рыб, и старших – менее упитанных, такой зависимости не наблюдается у рыб старших возрастов. Чаще они бывают менее упитанными.
Влияние вылова на стадо промысловой рыбы. Он оказывает самое разнообразное влияние. Изымая часть стада, вылов через разреживание популяции повышает обеспеченность пищей оставшейся части стада, что связано с изменением темпа роста особей, возраста достижения половой зрелости и предельного возраста. Таким образом, селективный вылов влияет на структуру популяции и даже других популяций, т.к. изымая часть стада, изменяет кормовую базу, создавая благоприятные условия для питания других видов рыб.
У интенсивно облавливаемых популяций промысловых рыб темп роста выше и популяционная плодовитость больше. Однако у всех популяций рыб при повышении интенсивности рыболовства положительные реакции наблюдаются только до определенного предела. При дальнейшем воздействии промысла регуляторные механизмы популяции нарушаются, и она перестает реагировать на дальнейшее разреживание стада, что является серьезным сигналом перелова.
Воспроизводство популяции не нарушается при определенной (для каждого вида) интенсивности вылова. Это происходит тогда, когда вылов изымает такую часть стада, к изъятию которой популяция приспособлена (выедание хищниками), когда вылов может быть компенсирован регуляторными механизмами популяции, т.е. когда вылов как бы является элементом среды вида. При подобной интенсивности вылова, и если не будут нарушены нерест и условия развития молоди, популяция может существовать многие годы.
У каждого вида в процессе его становления и приспособления к другим видам данного комплекса вырабатывается система приспособлений, защищающих его от хищников и система регуляторных механизмов, компенсирующих определенную смертность, т.е. в процессе исторического развития вид приспосабливается к определенной интенсивности смертности в каждой возрастной группе. Вылов является новым элементом среды, входящим в жизнь уже сформировавшегося вида.
У осетровых Азовского моря во время военного запуска рыболовства средние размеры и возраст осетровых в уловах значительно увеличились, а темп роста замедлился. Последующая интенсификация промысла привела к уменьшению средних размеров и возраста осетровых и усилению темпа их роста.
Интенсификация вылова пикши в предвоенные годы привела к разреживанию ее популяции и ускорению роста особей, это изменение темпа роста связано с более ранним созреванием состава нерестового стада. Наоборот, период запуска во время войны привел к постарению нерестовой популяции. С середины 50-х годов наблюдалось снижение уловов и ускорение роста рыб.
Сельдь. Раньше уловы сельди не определялись (в 1946 г. НЕАФК квотировали треску и камбалу)
В настоящее время в отношение некоторых стад сельди наблюдается длительная депрессия. Это свидетельствует о том, что интенсивность промысла этих стад достигла такой величины, которая не может быть компенсирована пополнением при обычных (средних) условиях воспроизводства. Это приводит к прогрессивному снижению численности стада. Причина не только в неурожайных годах (это низкое пополнение), но и влияет интенсивный вылов( на всю популяцию действует).
Изымая промысловых рыб промысел может приводить к замене ценных видов менее ценными (в заливе П. Великого желтоперая камбала была заменена на камбалу-ерша, лещ в озерах заменяется ершом и др.).
Далее, изымая часть популяции, промысел меняет отношение вида – жертвы с хищниками.
Таким образом, вылов изменяет взаимосвязи организмов в экосистеме водоема, иногда приводя к ее коренной перестройке.
Закономерности изменений структуры популяций под воздействием вылова. Вылов влияет на возраст, рост особей, соотношение полов и ряд других свойств популяции.
Теоретически селективным выловом можно изменять структуру популяции в очень значительной степени, даже довести ее до такого состояния, что популяция потеряет способность самовоспроизводиться. В 1919 году Ф. И. Баранов, исходя из теоретических представлений, что в водоеме молоди всегда достаточно, разработал концепцию воздействия промысла на стадо рыб.
1.“Кривая населения”, т.е. кривая численности рыб разного возраста в стаде, идентична кривой смертности.
2. Кривая улова отражает кривую распределения рыб по размерам и по возрасту в водоеме. Правая часть кривой улова подобна кривой населения, а левая есть результат отбора применяемой ячеей определенной величины.
3. Число ежегодно погибающих рыб промыслового размера равняется числу ежегодно подрастающих рыб минимальной длины независимо от вида кривой населения.
4. Для каждой популяции существует своя оптимальная допустимая величина улова. Поэтому основной метод регулирования рыболовства – это установление определенной нормы вылова.
5. Интенсивность лова влияет на форму кривой населения и коэффициент убыли. Она отражается на распределении в улове рыб разного возраста (длины и массы).
6. Кривая, отвечающая различной интенсивности убыли (годичная убыль), различна. Чем выше интенсивность промысла, тем круче кривая населения. Расчеты И.Ф. Баранова использовали Томпсон (1937), Э.С. Рессел (1939), И. Вестенберг (1957) и В.С. Ивлев (1959, 1961)
Таблица 1
Схема Томпсона
| Годовой класс | Запас, шт | Изъятие при 80% отлова, шт | Запас, шт | Изъятие при 50% отлова, шт |
| I | - | - | ||
| II | ||||
| III | ||||
| IV | ||||
| V | ||||
| VI | - | |||
| VII | - | - | ||
| VIII | - | - | ||
| IX | - | - | ||
| X | - | - | ||
| XI | - | - |
Из этой таблицы следует, что вылов изымает рыбу старших возрастов. Причина омоложения популяции не интенсификация промысла, а непропорционально большое пополнение, которое принимается одинаковым (1000 рыб) в случае изъятия и 50, и 80%.
Томпсон при определении оптимальной интенсивности рыболовства в приводимых примерах не учитывает ни естественной смертности рыб, ни индивидуального роста рыб.
Лекция№ 5.
Дата добавления: 2021-04-21; просмотров: 1192;
Поиск по сайту
Узнать еще
- Лексическая тема: Времена года
- Лекція 2. Тема: Новітні інформаційні технології та їх класифікація
- Лекція 3. Тема: Автоматизовані інформаційні технології, їх розвиток і класифікація
- Лекція 5. Тема: Організація аналітичного процесу
- Лекція 6. Тема: Документування та контроль у системі автоматизованого оброблення даних
- Лекція 7. Тема: Управлінські інформаційні системи аудиторської діяльності
- ЛЕКЦІЯ 7. Тема:Матеріально –технічна база ЦА. Основні техніко –економічні характеристики літаків.
- Материнская смертность

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
