ЭМБРИОНАЛЬНЫЙ, ПРЕДЛИЧИНОЧНЫЙ, ЛИЧИНОЧНЫЙ И МАЛЬКОВЫЙ ПЕРИОДЫ РАЗВИТИЯ РАСТИТЕЛЬНОЯДНЫХ РЫБ
В товарном рыбоводстве используют дальневосточных растительноядных рыб (обыкновенный, или белый толстолобик, пестрый толстолобик, белый амур), которые отличаются от остальных представителей карповых рыб тем, что у них жаберные перепонки сращены между собой, глаза располагаются очень низко, что отражено в латинском названии. Толстолобики достигают длины около 1 м и средней массы тела 12- 16 кг, а белый амур еще крупнее может достигать массы более 32 кг и длины тела более 1 м.
Обитают в реках Центрального и Южного Китая. Северная граница ареала проходит по реке Амур. Акклиматизированы в РФ - Краснодарском крае, Дагестане, государствах средней Азии и Украине. Белого амура вселяют в водоемы расположенные южнее 45° с.ш.
Все растительноядные рыбы – бысрорастущие, но более теплолюбивые, чем карп. В основном выращивают растительноядных рыб в прудовых хозяйствах в поликультуре с карпом, а также в озерах, водохранилищах и других неспускных водоемах. Для организации нагульных хозяйств на базе водохранилищ наиболее перспективны белый толстолобик и его гибриды с пестрым толстолобиком.
Белый толстолобик (Hypophthalmichtys molitrix Val.) питается фитопланктоном и детритом. Лучше всего потребляет диатомовые и зеленые водоросли и хуже синезеленые. Конкуренции в питании с карпом и другими видами в поликультуре практически нет.
Пестрый толстолобик (Aristichthys nobilis Rich.) наряду с зоопланктоном и детритом потребляет фитопланктон. При значительном увеличении плотности посадки может конкурировать с сеголетками карпа в питании зоопланктоном. В средней полосе растет лучше, чем белый толстолобик. В южных районах стран СНГ при хорошей обеспеченности кормом растет быстрее карпа.
Белый амур (Ctenopharyngodon idella Val.) питается высшей водной (нитевидный и узколистный рдест, роголистник, элодея, ряска) и наземной (злаки, клевер, люцерна, рогоз, тростник) растительностью. Способен быстро истребить собственную кормовую базу (особенно в южных районах). При недостатке растительности легко переходит на питание комбикормами, что может привести к конкуренции с карпом. При совместном выращивании белый амур имеет такой же темп роста, как пестрый толстолобик. Целесообразно использовать в прудовом хозяйстве в качестве биологического мелиоратора.
Кишечник растительноядных рыб приспособлен для переваривания такой низкокалорийной пищи как планктон. Так, его длина у белого толстолобика в 10-13 раз превышает длину тела.
Самки белого толстолобика созревают в южных районах, как правило в возрасте 3-4 лет, пестрого толстолобика - 4-5, белого амура – 4 лет. Самцы достигают половой зрелости на год раньше самок.
В природных условиях нерест растительноядных рыб происходит летом во время подъема воды в реках при температуре выше 20°С. Икра пелагическая, средняя плодовитость составляет 0,5 – 2 млн.шт. икринок. Белый толстолобик нерестится в верхних слоях воды, а пестрый в более нижней части пелагиали. Белый амур нерестится в русловых участках рек с турбулентным течением воды.
Перечисленные выше виды растительноядных рыб в прудах не размножаются и единственная возможность получения икры этих видов рыб – метод гипофизарных инъекций.
В биологии размножения и развития белого амура, белого и пестрого толстолобиков много общего, поэтому далее дается описание характера развития растительноядных видов рыб в ранний период жизни на примере белого амура.
Эмбриональный период развития растительноядных рыб
Этап 1. Оплодотворение, образование зиготы, образование перивителлинового пространства и образование бластодиска.
Стадия 1. Диаметр неоплодотворенной икринки 1,2-1,3 мм. Яйцевая оболочка плотно прилегает к поверхности икринки, она неклейкая и представлена первичной радиальной оболочкой. Икра прозрачная, бесцветная или слегка желтоватая (рис. 37, а).
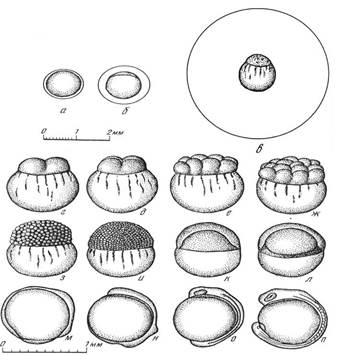
кп – купферов пузырек; ср – средний мозг; ск – слуховая капсула; хд – хорда; мт – миотом; ок – обонятельная капсула; жв – железа вылупления; жмш – желточный мешок
Рисунок 37 – Эмбриональный период развития растительноядных рыб на примере белого амура (обозначения в тексте)
Стадия 2. Возраст 10 мин после оплодотворения. Отделение яйцевой оболочки от желтка и концентрация плазмы на анимальном полюсе в виде прозрачной серповидной зоны (рис. 37, б)
Стадия 3. Возраст 40 мин. Происходит образование резко очерченного бластодиска. В основном завершается оводнение перивителлинового пространства. Диаметр икринки 3,8-4 мм, а собственно желтка 1,2-1,3 мм. Такое огромное перивителлиновое пространство уменьшает плотность икринки и обеспечивает ее плавучесть в потоках воды; в стоячей воде икринка опускается на дно и погибает (рис. 37, в)
Этап 2. Дробление от двух бластомеров до бластулы.
Стадия 4. Возраст 1 ч. На бластодиске закладывается первая борозда деления и образуются два бластомера (рис. 37, г).
Стадия 5. Возраст 1 ч 20 мин. Происходит закладка второй борозды деления и образование четырех бластомеров (рис. 37, д).
Стадия 6. Возраст 1 ч 40 мин. Отмечают закладку четвертой борозды деления и образование восьми бластомеров (рис. 37, е).
Стадия 7. Возраст 2 ч. На бластодиске образуются шестнадцать бластомеров (рис. 37, ж).
Стадия 8. Возраст 2 ч 30 мин. Деление идет как в экваториальном, так и в меридиональном направлениях. Образуется крупноклеточная морула (ранняя) (рис. 37, з).
Стадия 9. Возраст 4 ч 50 мин. Мелкоклеточная морула (поздняя) (рис. 37, и). Происходит завершение образования перивителлинового пространства. Окончательный диаметр оболочки варьирует от 4,32 до 5,32 мм.
Стадия 10. Возраст 6 ч. Отмечается начало образования бластулы (рис. 37, к).
Этап 3. Гаструляция- образование зародышевых пластов
Стадия 11. Возраст 7 ч 10 мин. Начинается обрастание бластодермой поверхности желтка (рис. 37, л ).
Стадия 12. Возраст 10 ч. Отмечается образование желточной пробки. Возраст 10 ч (рис. 37, м).
Стадия 13. Возраст 12 ч. 10 мин. Происходит замыкание желточной пробки. Зачаток тела приобретает вид утолщенного валика, расширенный головной отдел его начинается на анимальном полюсе и хвостовая часть его заканчивается на вегетативном полюсе (рис. 37, н).
Этап 4. Органогенез – дифференциация зародышевых пластов на зачатки основных органов.
Стадия 14. Возраст 15 ч. Происходит образование глазных пузырей, закладка хорды, начало сегментации мезодермы на сомиты. В головном отделе начинается закладка мозговых пузырей (рис. 37, о).
Стадия 15. Возраст 18 ч. Появление глазных бокалов и щелевидного углубления в зачатках глаз, продолжается сегментация тела. Хорда хорошо заметна (рис. 37, п).
Этап 5. Обособление хвостового отдела от желточного мешка, начало активного движения тела.
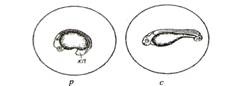
Стадии 16-18. Возраст 29-32 ч. Происходит выпрямление тела эмбриона. Начинаются энергичные колебательные и вращательные движения. Появление на голове и в области сердца желез вылупления (рис. 37, р, с).
Этап 6. Вылупление эмбриона из оболочки.
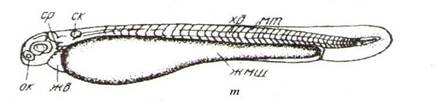
Стадия 19. Возраст 34 ч. Начинается вылупление с появления единичных предличинок (рис. 37, т).
Предличиночный период развития
Длина тела предличинки 5-5,2 мм. В туловищном отделе 29-31 сегмент, а в хвостовом – 12-14. Тело не пигментировано, окаймлено недифференцированной плавниковой складкой. В глазах черное пигментное пятнышко. Предличинки малоподвижны. В природных условиях пассивно сносятся течением в толще воды (рис. 37, т).
Этап 7. Образование эмбриональной сосудистой системы, начало кровообращения (рис. 38, а).
Стадия 20. Возраст - 51 ч. Длина - 6,5 мм. Формируются эмбриональные органы дыхания - хвостовая вена и кювьеровы протоки, расположенные на передней части желточного мешка. Движение предличинок пассивное. Питание эндогенное.
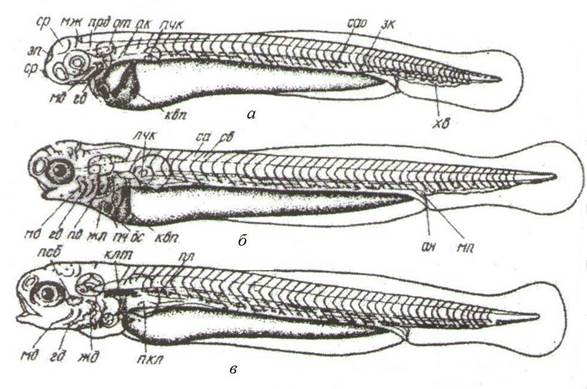
ан - анальное отверстие; жв - железа вылупления; жмш - желточный мешок; клт - клейтрум; кп- купферов пузырек; мж - мозжечок; мт - миотом; мп - мочевой пузырь; ок - обонятельная капсула; от - отолит; пл - плавательный пузырь; прд- продолговатый мозг; псб - псевдобранхия; пч- печень; пчк - зачаток предпочки; ск - слуховая капсула; ср- средний мозг; хд- хорда; эп- эпифиз; кс- кровеносная система: вс - венозный синус; сао – спинная аорта; зк – задняя кардинальная вена; мд – мандибулярная дуга аорты; гд – гиоидная дуга аорты; квп – кювьеров проток; хв – хвостовая вена; са - сегментальная артерия; св – сегментальная вена
Рисунок 38 – Предличиночный период развития растительноядных рыб
Этап 8. Образование и начало функционирования подвижного жаберно-челюстного аппарата (рис. 38, б, в).
Стадии 21-22. Возраст 76-96 ч. Длина 7,5 мм. Начало жаберного дыхания. Рот становится полуконечным и подвижным. Глаза полностью пигментированы. Предличинки становятся более подвижными. Питание эндогенное. Черные пигментные клетки (меланофоры) появляются на голове, над кишечником и в хвостовом отделе, на желточном мешке. Происходит редукция эмбриональных органов дыхания (кювьеровых протоков). Начинается закладка плавательного пузыря.
Личиночный период
Этап 1. Смешанное питание личинок.
Стадии 23-24. Возраст 4,5-6 сут. Длина 7,5-7,8 мм. Дыхание становится наружно-жаберным. Задняя камера плавательного пузыря заполняется воздухом. Личинка активна, заглатывает пищу, но продолжает также питаться за счет желточного мешка – питание смешанное. Пигментация тела усиливается. Личинки плавают в толще воды. На этом этапе рекомендуется перевозить личинок на дальние расстояния или высаживать в пруды (рис. 39, а, б).

Рисунок 39 - Личиночный период развития растительноядных рыб
Этап 2. Экзогенное питание личинок.
Стадия 25. Возраст 7 сут. Желточный мешок полностью резорбирован. Питается исключительно внешней пищей, в основном зоопланктоном. Происходит обособление лопастей непарных плавников. Жаберно-челюстной аппарат становится подвижным. Длина тела 7,6 мм (рис. 39, в).
Этап 3. Формирование непарных плавников (рис. 39, г, д, е).
Стадия 26. Возраст 9 сут. Длина 8 мм. Происходит образование лучей в нижней лопасти хвостового плавника. К возрасту 16-ти суток все непарные плавники имеют плавниковые лучи.
Стадии 27-28. Конец хорды загнут кверху. В хвостовом плавнике образуется выемка. Заполняется воздухом передний отдел плавательного пузыря. Закладываются брюшные плавники.
Этап 4. Формирование парных плавников.
Стадии 29-30. Возраст 20-22 сут. Длина тела – 14,2 мм. В парных плавниках образуются плавниковые лучи. Сохраняется преанальная плавниковая складка (рис. 39, ж).
Мальковый период развития
Этап 1. Начало закладки чешуи.
Возраст 1 мес. Длина тела 2 см. Вдоль боковой линии начинает появляться чешуя. Происходит переход на питание высшей водной растительностью (рис. 40, а).
Этап 2. Окончание формирования чешуйного покрова.
Возраст 1,5 мес. Длина тела 4-5 см. Преанальная складка исчезла. Тело полностью покрыто чешуей. Видны отверстия канала боковой линии (рис. 40, б).
Рисунок 40 – Мальковый период развития растительноядных рыб
Не смотря на общую схему развития, в эмбриональном и постэмбриональных периодах существуют различия по морфологическим и морфометрическим показателям у растительноядных рыб (табл. 3).
Аномалии развития.При искусственном разведении рыб не всегда удается обеспечить оптимальные условия для нормального протекания физиологических процессов и правильного развития эмбрионов и личинок. Это ухудшает рыбоводные качества икры, вызывает разнообразные нарушения в строении эмбрионов и снижает их жизнеспособность.
Таблица 3 - Различия в развитии белого амура, белого и пестрого толстолобиков
| Морфологические и морфометрические признаки | Белый амур | Белый толстолобик | Пестрый толстолобик | ||||
| Икра (набухшая) | |||||||
| Средний диаметр, мм оболочки желточного мешка | 4,38-5,22 1,21-1,36 | 3,80-4,50 1,10-1,20 | 4,82-5,63 1,42-1,50 | ||||
| Предличинки | |||||||
| Количество миотомов в туловище в хвосте | 29-31 12-14 | 24-26 14-17 | 24-26 14-17 | ||||
| Отношение длины туловища к длине хвоста | 2,5 | 2,0 | 2,0 | ||||
| Наличие черного пигмента на желточном мешке | Пигмент только спереди | Пигмент спереди и на брюшной части | Пигмент спереди и слабо на брюшной части | ||||
| Личинки | |||||||
| Количество миотомов в туловище в хвосте | 29-31 12-14 | 24-26 14-17 | 24-26 14-17 | ||||
| Отношение длины туловища к длине хвоста | 2,5 | 2,0 | 2,0 | ||||
| Пигментация преанальной плавниковой складки | Пигмент отсутствует | Пигмент развит сильно | Пигмент развит несколько слабее | ||||
| Соотношение развития челюстей, начиная с третьего этапа | Челюсти оканчиваются на одном уровне | Челюсти оканчиваются на одном уровне | Нижняя челюсть выступает вперед верхней | ||||
| Развитие грудных плавников на последнем 4-м этапе | Далеко не доходят до основания брюшных плавников | Далеко не доходят до основания брюшных плавников | Заходят за основание брюшных плавников | ||||
| Развитие спинного плавника | Оканчивается, не доходя до уровня начала основания анального плавника | Оканчивается на уровне начала основания анального плавника | Оканчивается на уровне середины или конца основания анального плавника | ||||
| Мальки | |||||||
| Соотношение развития челюстей | Как у личинок | ||||||
| Развитие грудных плавников | То же | ||||||
| Развитие спинного плавника | « » | ||||||
| Число лучей в анальном плавнике | 8-9 | 11-14 | 11-14 | ||||
| Чешуйный покров | Чешуя крупная | Чешуя мелкая | Чешуя мелкая | ||||
| Развитие брюшного киля | Киль отсутствует | Киль развит от основания грудных плавников до ануса | Киль развит лишь от основания брюшных плавников до ануса | ||||
Наиболее частой причиной неудовлетворительного качества икры и уродства личинок является передержка икры в теле самки после ее овуляции. Надежным критерием качества икры служит процент ее оплодотворения и морфологическая картина развития. Доброкачественная икра имеет высокий процент оплодотворения (90-95 %, а иногда 100 %) и не имеет нарушений в развитии.
У недоброкачественной икры нередко наблюдается высокий процент оплодотворения, но развитие идет ненормально. Поэтому один лишь показатель процента оплодотворения икры не может служить критерием качества. Необходимо контролировать процесс на различных этапах развития. На каждом этапе развития могут быть обнаружены характерные аномалии, зависящие как от качества икры, так и от условий среды.
Например, наблюдаются аномалии набухания, причиной которых является разноразмерность икринок, полученных от одной самки; такая икра, как правило, хорошо оплодотворяется, но имеет большой отход в период инкубации и дает значительное количество уродливых личинок.
Аномалии оболочки икры наблюдаются при склеивании икринок. Это происходит в том случае, если икру в момент приливания воды после осеменения тщательно не перемешивать. В месте склеивания с другой икринкой наружный слой оболочки разрывается и в разрыв выпячивается внутренний слой.
При аномалии дробления бластодиска наиболее частым нарушением является отрыв бластомеров и различная величина бластомеров.
При аномалии желточного мешка, желток у недоброкачественной икры по сравнению с доброкачественной имеет более крупные и неоднородные гранулы. Эмбрионы с такими нарушениями желтка обычно доживают до вылупления, но при переходе на предличиночный период развития оказываются нежизнеспособными и погибают в значительных количествах.
Ложное развитие неоплодотворенной икры происходит своеобразно. Попадая в воду, она набухает, на анимальном полюсе образуется плазменный бугорок, который начинает дробиться. Однако дробление оказывается ложным, так как деление бластомеров не доходит до конца, образуются разноразмерные, асимметрично расположенные псевдобластомеры, представляющие собой безъядерные выпячивания цитоплазмы.
Во время такого беспорядочного псевдодробления неоплодотворенная икра становится хорошо отличимой от оплодотворенной, бластомеры которой имеют одинаковые размеры и четкие контуры. Поэтому наиболее пригодными для определения процента оплодотворения являются стадии дробления от 4-8 бластомеров до ранней морулы. Позднее границы ложных бластомеров исчезают, цитоплазма приобретает ровную поверхность и начинает совершать ложную гаструляцию.
Псевдогаструляция завершается разрушением поверхности плазменного слоя и коагуляцией вытекающего из разрыва желтка.
Массовая гибель неоплодотворенной икры совпадает с периодом начала формирования тела зародыша оплодотворенной икры.
Водянка зародышей чаще проявляется после начала образования сердца. Эта аномалия заключается в чрезмерном увеличении и оводнении околосердечной полости. Высокая степень водянки приводит к значительной деформации сердца.
Водянка может образоваться позади околосердечной полости, под передним или задним отделом кишечника, в мочевом пузыре и др. Жизнеспособными бывают лишь зародыши с очень слабо выраженными признаками водянки.
Деформации тела зародыша - искривление туловища, хвостового отдела, диспропорции отдельных частей тела наблюдаются обычно при водянках. Причины подобных аномалий - резко выраженная недоброкачественность икры, нарушение условий инкубации, в частности понижение температуры воды и других условий.
Дата добавления: 2016-08-06; просмотров: 9173;
Поиск по сайту
Узнать еще
- Filariidae:. Onchocerca volvulus Систематика, морфология, цикл развития, патогенное действие, диагностика и профилактика онхоцеркоза и лоаоза.
- II. Завоевание Китая маньчжурами. Экономическое положение страны в XVII – начале XIX вв.: аграрная политика Цинской династии, особенности развития городского ремесла
- II. Особенности развития турецкой буржуазии. Становление младотурецкого движения
- III Всебелорусское собрание. Программа социально-экономического развития Республики Беларусь на 2006 – 2010 гг.
- III. Периоды колонизации стран Востока
- IV. Внешние условия развития отрасли информационных технологий
- VII. Пути развития национального экологического законодательства с учётом международных обязательств РК
- а) Краткая характеристика культурного развития.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
