Немитохондриальные электрон-транспортные цепи растительной клетки
У растений, как и у других организмов, имеются системы окисления восстановленных пиридин- и флавиннуклеотидов не только с помощью терминальных оксидаз митохондрий, но и в редокс-цепях эндоплазматического ретикулума, плазмалеммы и цитоплазмы. В процессах окисления, катализируемых немитохондриальными оксидазами, не происходит образование АТР. К таким оксидазам относятся, например, Сu-содержащие ферменты – полифенолоксидаза и аскорбатоксидаза. Медьсодержащие оксидазы способны восстанавливать оба атома молекулярного кислорода до Н2О, чем напоминают цитохром с оксидазный комплекс, который также содержит медь. Однако они не содержат железа.
Для окисления аскорбиновой кислоты в дегидроаскорбиновую в растительных клетках имеется специальная аскорбатоксидазная система, способная осуществлять такое превращение. Функционирование этой системы обеспечивает окисление таких субстратов, как НАД(Ф)Н, восстановленная форма глутатиона и аскорбиновая кислота.
Полифенолоксидаза окисляет фенольные соединения (моно-, ди- и трифенолы) с образованием соответствующих хинонов. Именно активностью полифенолоксидазы объясняется потемнение поверхности разрезанного яблока или картофельного клубня, поскольку при ранениях растительных тканей этот фермент превращает фенолы в токсичные для патогенов хиноны, что препятствует распространению инфекции. Полифенолоксидаза участвует в окислении полифенолов и дубильных веществ, происходящем при скручивании и завяливании чайного листа, ее действием объясняется также потемнение плодов при сушке. Активность полифенолоксидазы возрастает в период старения тканей.
Свойство аутооксидабельности, т. е. прямого окисления кислородом, присуще лишь немногим органическим соединениям. Восстановленные флавины доступны быстрому прямому окислению молекулярным О2. НАДН и НАДФН самопроизвольно кислородом не окисляются. «Самоокисляемость» позволяет флавинам некоторых ферментов переносить электроны непосредственно к О2, а также создает основу для функционирования флавинов в реакциях гидроксилирования. В цитоплазме растительных клеток функционируют флавопротеиновые оксидазы, которые реагируют с О2, образуя Н2О2.
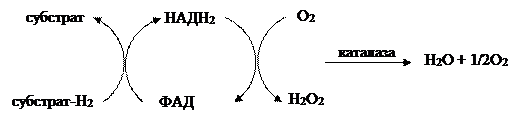 |
Системы терминальных оксидаз в растениях (цитохромоксидаза, альтернативная оксидаза, аскорбатоксидаза, полифенолоксидаза, флавопротеиноксидаза) различаются по чувствительности к изменению парциального давления кислорода и температуры. Комплекс цитохром с оксидазы обладает высоким сродством к О2 и поэтому активно функционирует при низком парциальном давлении кислорода. Однако его активность сильно меняется при колебаниях температуры. Флавопротеиноксидазы меньше зависят от температуры, но имеют более низкое сродство к О2. Полифенолоксидаза и аскорбатоксидаза занимают по этим характеристикам промежуточное положение.
В мембранах эндоплазматического ретикулума растительных клеток найдены электрон-транспортные цепи, в которых проявляется активность монооксигеназ. Используя восстановители (НАДН или НАДФН) и цитохром b5 , они осуществляют реакцию десатурации– образования ненасыщенных жирных кислот из насыщенных:
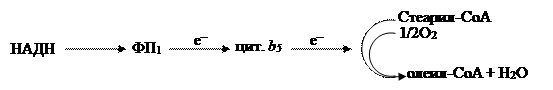 |
В другой редокс-цепи цитохром Р450, используя кислород, осуществляет реакцию гидроксилирования различных соединений:
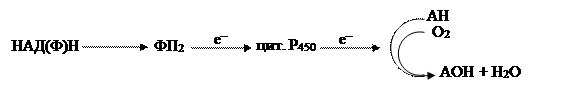 |
Процесс гидроксилирования необходим для синтеза некоторых соединений, детоксикации ряда вредных метаболитов, которые теряют свою активность, и, наконец, для превращения нерастворимых в воде веществ в растворимые.
В плазматической мембране есть редокс-цепь, которая окисляет НАДФH и передает электроны через цитохром b559, превращая его в супероксид-радикал. Эта НАДФH-оксидаза играет защитную роль в реакции сверхчувствительности при повреждении растительных клеток патогенами.
Дата добавления: 2016-07-11; просмотров: 1866;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. Поверхностный аппарат клетки.
- III. Расчёт электрического состояния цепи с последовательным соединением элементов L, R, C.
- IV. Расчёт цепи с параллельным соединением R, L, C элементов
- RLC-контур с параллельным соединением элементов в цепи переменного тока.
- А) Определение резистентности грудной клетки
- Активное сопротивление, индуктивность и емкость в цепи переменного тока.
- Алгебраическая сумма токов, текущих в участках цепи, соединённых в

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
