Цианидустойчивое дыхание растений
Существенное отличие растительных митохондрий заключается также в том, что во внутренней мембране наряду с широко распространенным цитохромным путем переноса электронов по ЭТЦ может функционировать альтернативный, устойчивый к действию цианида транспорт электронов. В этом случае поток электронов обходит комплексы III и IV переносчиков ЭТЦ, где транспорт электронов сопряжен с активным транспортом протонов и фосфорилированием. В результате отношения Ф : О и, следовательно, эффективность дыхания значительно снижаются. Альтернативный путь характерен практически для всех видов растений.
Такая особенность дыхания растений объясняется тем, что во внутренней мембране растительных митохондрий функционирует альтернативная цианидустойчивая оксидаза, которая обеспечивает передачу электронов от убихинона на кислород с образованием Н2О. Этот процесс специфически угнетается рядом соединений, к которым относится, например, салицилгидроксамовая кислота. Когда электроны от убихинона попадают на кислород, то пропускается два участка электрон-транспортной цепи (комплексы III и IV), на которых была бы возможна генерация энергии (вначале в виде протонного градиента, а затем и АТФ). Поэтому происходит рассеивание энергии в виде тепла. Чувствительность альтернативной цианидустойчивой оксидазы к кислороду несколько ниже, чем у цитохромоксидазы.
Альтернативная оксидаза представляет собой интегральный мембранный белок, состоящий из двух идентичных субъединиц, молекулярная масса каждой из которых составляет 35,5 и 37,0 кДа. Регулирование активности этого фермента осуществляется путем образования (димерная форма) или расщепления (мономерная форма) дисульфидного мостика между мономерами:
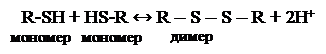 |
У многих цветков перед образованием пыльцы наблюдается быстрое повышение активности этого фермента. Это приводит к резкому возрастанию температуры тканей цветка, которая, например у Sauromatum guttаtum достигает 25 оС. Одновременно активируется синтез салициловой кислоты, а также определенных аминов и индольных соединений, которые служат аттрактантами для насекомых-опылителей. Такие условия могут поддерживаться до 7 ч. Повышение температуры характерно для соцветий и цветков многих видов ароидных.
Физиологическая роль этого пути не совсем ясна. По мнению Г. Ламберса (1985 г.) альтернативный путь служит в качестве «клапана» сброса избытка энергии в растении, особенно для тех органов, куда поступают ассимилянты с высоким отношением С:N и где невозможно хранить избыточное количество углеводов, например в корнях. Альтернативный путь действует тогда, когда поступление ассимилянтов превышает спрос и скорость переноса электронов на убихинон превосходит способность цитохромного пути транспорта электронов или последний насыщен.
Однако оценить вклад альтернативного пути в общее дыхание сложно, поскольку установлены факты переключения переноса электронов с альтернативного на цитохромный путь и обратно.
Считается также, что альтернативная цианидустойчивая оксидаза активируется при стрессовых воздействиях, которые повреждают работу основной электрон-транспортной цепи митохондрий.
Еще характерной особенностью митохондрий растительных клеток, в отличие от животных, является наличие маликэнзима, который катализирует окислительное декарбоксилирование малата:
 |
Пируват далее окисляется в цикле Кребса, а НАДН – в электрон-транспортной цепи. Таким образом, растительные митохондрии способны окислять через цикл Кребса не только образующийся в гликолизе пируват, но и транспортируемый из цитоплазмы малат.
Дата добавления: 2016-07-11; просмотров: 1658;
Поиск по сайту
Узнать еще
- Адаптация растений и животных к световому режиму.
- Анатомическое строение стебля однодольных и двудольных растений. Строение стебля двудольных и голосеменных древесных растений. Структура древесины
- Анаэробное окисление: нитратное и сульфатное дыхание
- Ассортимент растений, рекомендуемый для создания санитарно-защитных зон и озеленения города
- Б) Адаптации растений к тепловому режиму
- Б) Экологические группы растений по отношению к свету
- Бактериальный фотосинтез и его типы. Отличие бактериального фотосинтеза от фотосинтеза растений.
- Банк in vitro и криоконсервация; их значение для сохранения генофонда растений

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
