Охарактеризовать формы размножения бурых водорослей.
3. Какие принципы лежат в основе классификации водорослей данного отдела?
4. Для каких форм бурых водорослей характерно наибольшее анатомическое и морфологическое расчленение таллома?
5. Как осуществляется смена ядерных фаз и форм развития в различных классах бурых водорослей (на примере эктокарпуса, диктиоты и ламинарии)?
6. Каковы цикл развития и место редукционного деления ядра у фукуса?
7. Каковы филогенетические связи бурых водорослей с другими представителями слоевищных растений?
8. Каково значение бурых водорослей в природе и народном хозяйстве?
1.12. ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ, ИЛИ БАГРЯНКИ (RHODOPHYTA)
Представители отдела в подавляющем большинстве — многоклеточные организмы сложного морфологического и анатомического строения, и только немногие, наиболее примитивные, имеют одноклеточное или колониальное слоевище кокко- идной структуры. Некоторые багрянки — крупные водоросли, достигающие в длину от нескольких сантиметров до 2 м, но среди них немало и микроскопических.
По форме красные водоросли бывают в виде нитей, кустиков, пластинок, пузырей, корок, кораллов и т.д. Большого разнообразия достигают пластинчатые формы. Встречаются пластины цельные и сложно рассеченные, с дополнительными выростами по краю и на поверхности. Некоторые багрянки сильно кальцинированы и напоминают окаменелости.
При всем внешнем многообразии красным водорослям свойствен единый план строения слоевища - в его основе у всех многоклеточных багрянок лежит гетеротрихальная структура.
Ветви красных водорослей делятся на две категории. Одни — основные длинные ветви, которые растут в течение всего периода роста водоросли, так называемые ветви неограниченного роста. Другие — растут только до определенного предела и всегда остаются более или менее короткими — ветви ограниченного роста.
Кроме того, существуют и специализированные ветви, выполняющие роль усиков, или ризоидов, служащих для дополнительного прикрепления либо сцепления друг с другом.
Паренхиматозный тип организации фактически отсутствует. Единственный пример такого слоевища — представитель класса Бангиевые (порфира). У большинства же багрянковых слоевища псевдопаренхиматозного типа (за счет переплетения ветвей одной оси — одноосевое строение или многих — многоосевое). Увеличение размеров слоевищ у примитивных форм осуществляется за счет диффузного деления клеток, у более организованных — в результате деления верхушечных клеток, а у ряда видов — за счет верхушечной или краевой меристемы. Органами прикрепления к субстрату служат ризоиды, присоски, подошвы или стелющиеся ризоидальные пластины.
Клетки красных водорослей покрыты оболочкой, в которой различимы внутренний, целлюлозный, и наружный, пектиновый, ослизняющийся слои. Получаемый из последнего агар- агар содержит кроме пектина сахара и белки. Оболочка может быть пропитана известью, солями магния или железа.
Цитоплазма отличается повышенной вязкостью, плотно прилегает к стенкам, чувствительна к изменению солености среды. У высокоорганизованных водорослей клетки многоядерные, у менее организованных — одноядерные.
Форма хроматофоров зависит от интенсивности освещения, размеров и возраста клеток. Однако чем выше организация водоросли, тем больше в ее клетках хроматофоров и тем постояннее их форма (преимущественно линзовидная).
Пиреноиды у многих видов отсутствуют. Как и у других водорослей, окраска пластид и всего тела красной водоросли обусловлена сочетанием нескольких пигментов: хлорофиллов а и d, фикобилинов (фикоцианин, фикоэритрин, аллофикоци- анин) и каротиноидов. Окраска таллома варьирует от малиново-красной (преобладание фикоэритрина) до голубовато-сталь- ной (при избытке фикоцианина).
На примере водорослей отчетливо прослеживается приспособительный характер пигментного аппарата у водных растений. Известно, что при прохождении света через толщу воды отсекается оранжево-красная часть спектра и до больших глу бин доходят в основном зеленые, голубые и синие лучи. Соответственно у растений на мелководье преобладают зеленые пигменты, у более глубокообитающих водорослей (бурые) к ним добавляются бурые пигменты (фукоксантин и др.), а у самых глубоководных красных — красные пигменты (фикоэри- трин и др.). Помимо того, проникновение красных водорослей на значительные глубины можно объяснить их способностью (благодаря красным пигментам) улавливать даже незначительное количество света.
Запасные продукты представлены в виде флоридозида, содержащего галактозу и глицерин, сахарозы, жиров и «багрянкового крахмала», зерна которого откладываются в цитоплазме, а не в хроматофорах.
Багрянки обладают сложным, не встречающимся у других водорослей циклом развития, своеобразным строением женского органа размножения — карпогона и сложными процессами развития зиготы. Подвижные стадии в цикле развития отсутствуют, их споры и гаметы лишены жгутиков.
Способы размножения красных водорослей весьма многообразны. Вегетативное размножение свойственно лишь примитивным. Оно осуществляется за счет образования дополнительных побегов, отрастания нового таллома от подошвы старого, отмершего, а также путем деления клеток. Оторванные участки талломов погибают.
Собственно бесполое размножение осуществляется моно-, би-, тетра- и полиспорами, образовавшимися в спорангиях. Тетраспоры формируются на диплоидных бесполых организмах — спорофитах (тетраспорофитах). В тетраспорангиях перед образованием тетраспор происходит мейоз.
Половой процесс оогамный. Карпогон обычно состоит из расширенной базальной части — брюшка (с ядром внутри) и трубчатого выроста - трихогины, принимающей спермации. Сперматангии — небольшие бесцветные клетки, содержимое которых освобождается в виде мелких, голых, лишенных жгутиков мужских гамет — спермациев. Оплодотворение яйцеклетки осуществляется за счет перемещения ядра спермация по трихогине в карпогон. После оплодотворения базальная часть карпогона отделяется перегородкой от трихогины, которая отмирает и претерпевает дальнейшее развитие, приводящее к образованию карпоспор. Детали этого развития имеют важное значение при классификации багрянок. У одних красных водорослей содержимое зиготы делится с образованием непо движных голых спор — карпоспор, у других из оплодотворенного карпогона образуется система специальных диплоидных нитей — гонимобластов, клетки которых превращаются в кар поспорангии, производящие по одной диплоидной карпоспо- ре. У большинства багрянок развитие карпоспор проходит с участием ауксилярных клеток. В таких случаях гонимобласт развивается не из брюшка карпогона, а из ауксилярной клетки. Если же ауксилярные клетки удалены от карпогона, из его брюшка после оплодотворения вырастают соединительные (ообластемные) нити; клетки их диплоидны. Ообластемные нити подрастают к ауксилярным клеткам и в точке их соприкосновения оболочки растворяются, после чего происходит плазмогамия, в результате развивается гонимобласт с карпо- спорами — карпоспорофит. Следовательно, ауксилярные клетки выполняют вспомогательную функцию — стимулируют деление ядра клетки соединительной нити и поставляют дополнительное питание. У наиболее высокоорганизованных красных водорослей (флоридеофициевые) ауксилярные клетки развиваются после оплодотворения карпогона в непосредственной от него близости. Ообластемные нити у этих водорослей не образуются. Ауксилярная клетка, находясь рядом с брюшком карпогона, сливается с ним и образует прокаргшй, из которого развивается гонимобласт с карпоспорами. Карпо- спорангии часто располагаются тесными группами — цисто- карпиями, которые у многих представителей одеты псевдопаренхиматозной оболочкой, развивающейся из соседних с кар погоном клеток.
Циклы развития красных водорослей разнообразны. У одних представителей флоридеофициевых происходит смена трех форм развития: гаплоидный гаметофит, диплоидные карпо- и тетраспорофит. В этом случае зигота делится без редукции числа хромосом, формируя диплоидный карпоспорофит (гонимобласт с карпоспорами). Карпоспоры прорастают с образованием диплоидного нитчатого тетраспорофита, на котором в результате мейоза образуются гаплоидные тетра- споры, дающие начало гаметофитам. Таким образом, имеются 2 свободноживущие формы одной и той же водоросли — тетраспорофит и гаметофит.
У других водорослей (с гетероморфной сменой форм развития) часто бывает слабо развит и даже редуцирован тетра- и карпоспорофит, иногда редуцирован гаметофит (он формируется на спорофите).
Красные водоросли — типичные морские организмы. В пресных водоемах обитает лишь небольшое количество видов.
Принцип, положенный в основу классификации багрянок, - строение женских генеративных органов и процесс развития гонимобласта. По этой же системе все красные водоросли делятся на 2 класса — Бангиофициевые и Флоридеофициевые.
Класс Бангиофициевые (Вangiophyceae)
Класс Бангиофициевые объединяет одноклеточные, колониальные и многоклеточные (паренхимного, реже — одноряд- но-нитчатого строения), прикрепленные к субстрату водоросли. Рост слоевища у них диффузный, т.е. за счет деления всех клеток. Клетки одноядерные, с одним хроматофором звездчатой формы и пиреноидом.
Бесполое размножение осуществляется делением клеток. У некоторых примитивных видов этот способ размножения является единственным. Бесполое размножение осуществляется также моноспорами.
Половой процесс известен у немногих видов. Карпогон без трихогины. Зигота прорастает непосредственно группой карпоспор. Некоторым высокоорганизованным представителям свойственна смена половой и бесполой (диплогаплофазный цикл) форм развития.
Бангиофициевые — в основном пресноводные и наземные обитатели. Морские формы распространены в прибрежной полосе всех морей, но в умеренных широтах представлены богаче, чем в тропиках. Класс Бангиофициевые включает 70 видов из 30 родов. Наиболее известен порядок Бангиевые.
Порядок Бангиевые (Bangiaks). Включает виды, имеющие многоклеточные с интеркалярным ростом слоевища нитчатой и пластинчатой формы. Все клетки однотипные, не дифференцированные на осевые и коровые, одеты оболочкой, соединены между собой посредством сочленяющих пор.
Бесполое размножение осуществляется посредством моно- и полиспор; половой процесс оогамный. После оплодотворения содержимое карпогона делится с образованием 4 — 64 карпоспор. Большинство представителей обитает в морях, но есть и пресноводные (например, бангия).
Род порфира (Porphyra) (рис. 1.33) имеет вид пластинки розовато-пурпурного цвета с гладкими или волнистыми края-
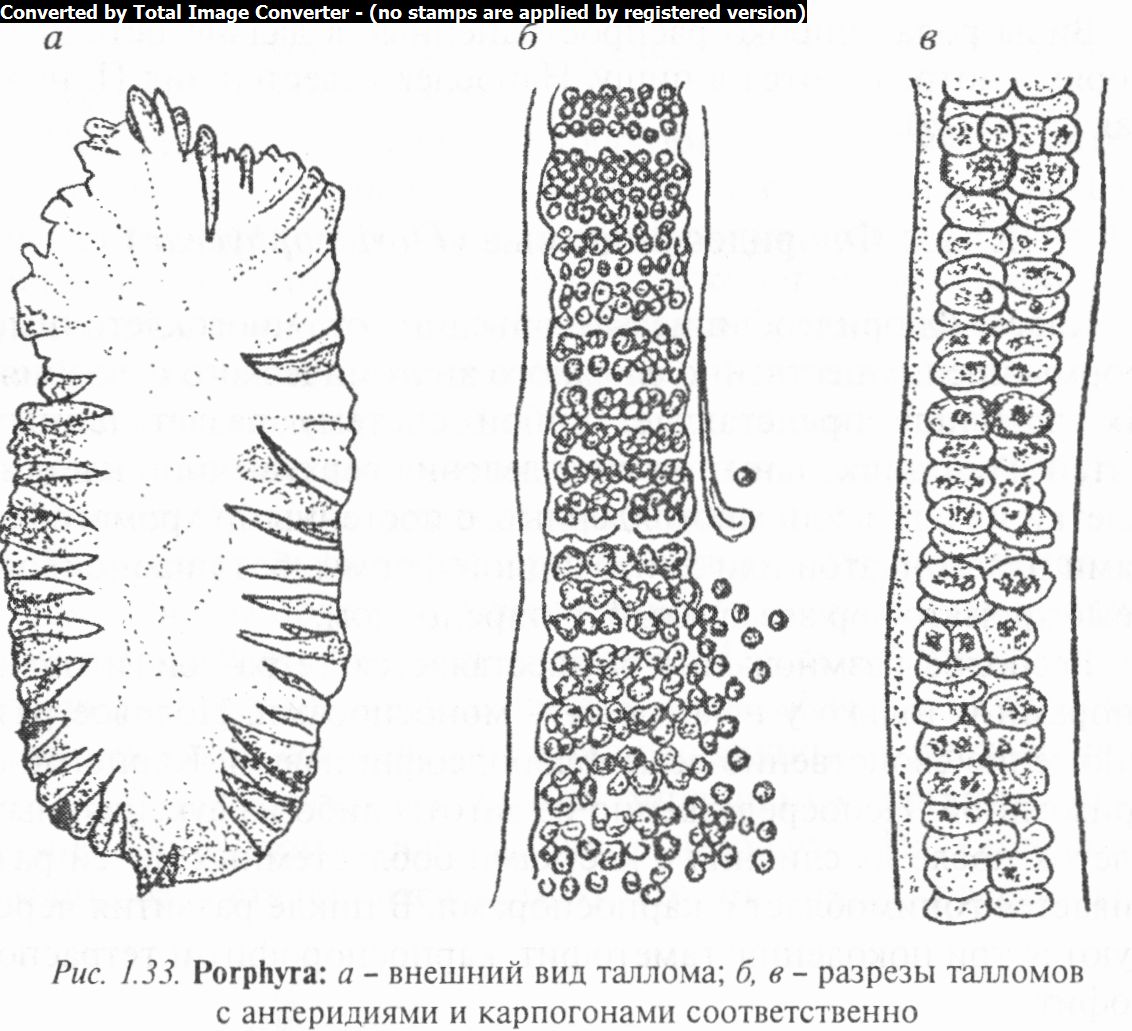
ми величиной до 50 см (иногда до 2 м) в длину и до 10 см и более в ширину. Пластинка состоит из одного или двух слоев клеток и при помощи короткого черешка и подошвы прикрепляется к подводным предметам.
Из клеток таллома формируются органы полового размножения: мужские (сперматангии) и женские (карпогоны). Спер- матангии — мелкие клетки, образующиеся за счет взаимно перпендикулярных делений клеток таллома и дающие по одному спермацию. Карпогоны лишь незначительно отличаются от вегетативных клеток наличием небольшого сосочковидного выроста. Типичная трихогина отсутствует. После оплодотворения в карпогоне образуется 4-64 карпоспоры. Судьба прорастающих карпоспор различна. При оптимальных условиях они снова дают диплоидные пластинчатые талломы. В других условиях карпоспоры прорастают в однорядно-нитчатые разветвленные спорофиты, отдельные клетки которых функционируют как моноспорангий. В них образуются гаплоидные моноспоры, дающие при прорастании пластинчатые слоевища — гаметофиты. Таким образом, у порфиры имеется чередование гамето- и спорофита — гетероморфная смена форм развития.
Виды рода, широко распространенные в дальневосточных морях, употребляются в пищу. Наиболее известен вид П. нежная (P. tenera).
Класс Флоридеофициевые (Florideophyceae)
Класс Флоридеофициевые объединяют многоклеточные формы преимущественно сложного анатомического строения. Их слоевище представляет собой систему разветвленных нитей. Рост апикальный за счет деления верхушечной клетки. Клетки их одно- или многоядерные, с постенными хроматофо- рами пластинчатой или линзовидной формы, без пиреноидов; реже хроматофор звездчатый, с пиреноидом.
Бесполое размножение осуществляется тетра-, би- и полиспорами и только у некоторых — моноспорами. Половое размножение свойственно всем флоридеофициевым. Карпогон с трихогиной. Непосредственно из зиготы либо из ауксилярных клеток после их слияния с клетками ообластемных нитей развивается гонимобласт с карпоспорами. В цикле развития чередуются три поколения: гаметофит, карпоспорофит и тетраспорофит.
Подавляющее большинство представителей класса — типичные морские обитатели, распространенные во всех морях земного шара.
Класс Флоридеофициевые делят на 6 порядков на основании особенностей морфологического и анатомического строения водорослей, деталей развития зиготы и строения аукси- лярной системы.
Порядок Немалиальные (Nemaliales). Включает водоросли одно- или многоосевого строения. Для них характерно отсутствие ауксилярных клеток; гонимобласты развиваются непосредственно из оплодотворенного карпогона или из его дочерней клетки. Бесполое размножение совершается преимущественно моноспорами. В цикле развития обычно наблюдается гетероморфная смена трех форм развития.
Виды порядка живут главным образом в морях, но встречаются и в пресных водах.
Род батрахоспермум (Batrachospermum) объединяет виды, имеющие форму рыхлого слизистого цилиндра или нити с мутовчато расположенными боковыми веточками ограниченного роста, делающими их похожими на четки или небольшие (от 3 до 8 см) кустики. Таллом бывает оливково-зеленого, буроватого или стального цвета.
Центральная ось слоевища образована длинными бесцветными клетками, расположенными в один ряд. От границы двух соседних клеток берут начало мутовки боковых разветвлений, обильно ветвящихся и сложенных из коротких бочонкообраз- ных клеток с многочисленными дисковидными хроматофора- ми. Это так называемые ассимиляторы. Крупные клетки основных ветвей в более старых частях таллома покрыты многоклеточными нитями, которые могут переплетаться и у некоторых видов образовывать обертку (своеобразную кору).
Размножается батрахоспермум главным образом половым путем. Органы полового размножения (карпогон и сперматан- гии) формируются на ассимиляторах. После оплодотворения из брюшка карпогона развивается гонимобласт с округлыми карпоспорами, собранными в тесную группу, — цистокарпий (рис. 1.34), являющийся новым диплоидным организмом — карпоспорофитом, живущим на гаметофите. Из карпоспор развиваются стелющиеся нити, от которых отходят вертикаль-
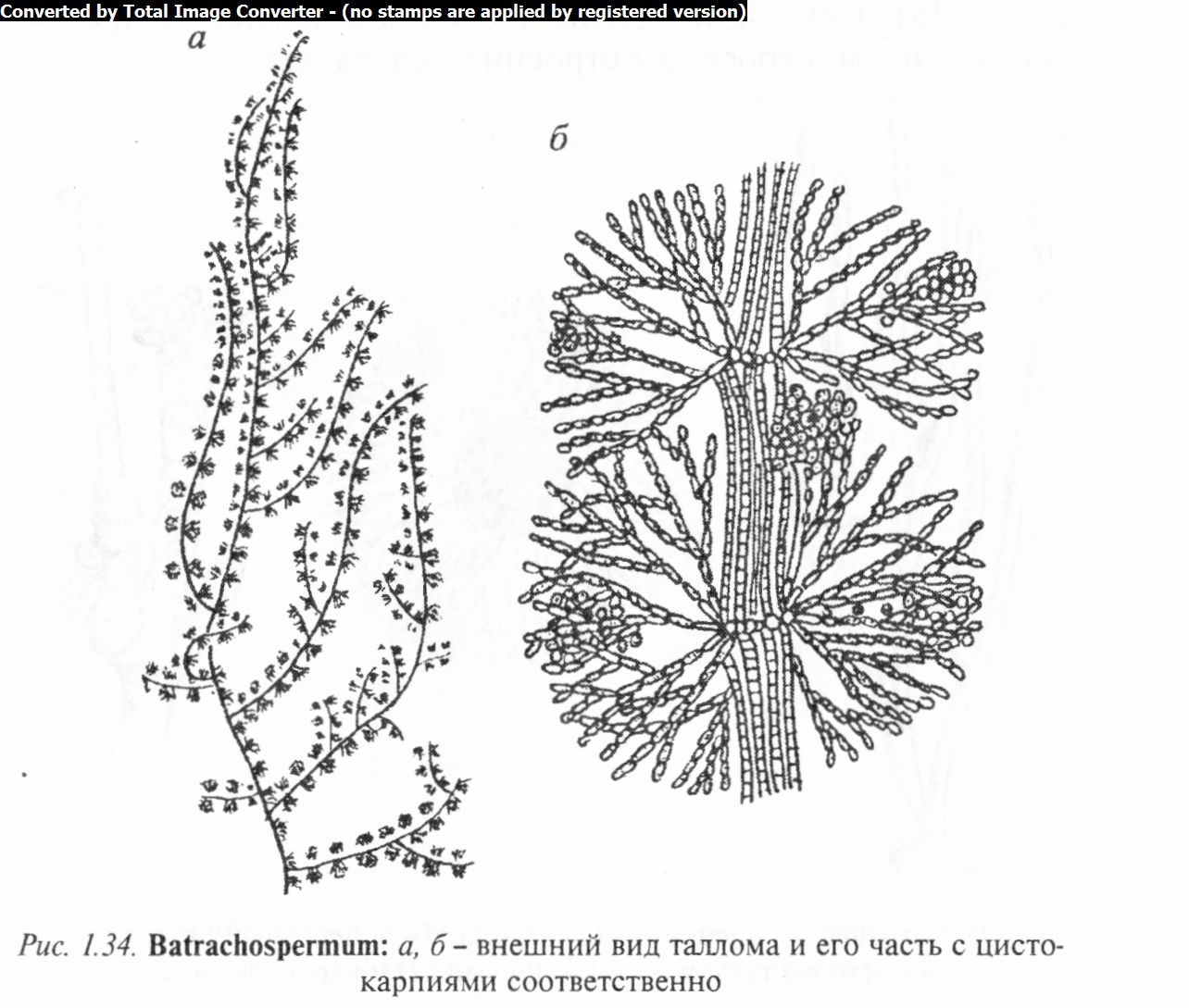
ные ветвящиеся нити совсем иного внешнего вида, чем таллом батрахоспермума. Эту стадию ранее считали самостоятельной водорослью и описывали под родовым названием «шантран- зия» (Chantransia). Она способна размножаться моноспорами. Из верхушечных клеток нитей данной стадии могут развиваться типичные талломы батрахоспермума. Одни исследователи предполагают, что шантранзия диплоидна и является спорофитом, другие рассматривают ее как неполовозрелую стадию батрахоспермума.
Известно 100 видов рода, распространенных во всех частях света. Встречаются, как правило, в богатой кислородом воде, отдельные — на торфяных болотах. Типичным и распространенным видом является Б. четковидный (В. moniliforme), занесенный в Красную книгу Республики Беларусь.
Увидов рода немалион (Nemalion) слизистое шнурообразное слоевище, состоящее из пучка клеточных нитей, каждая из которых растет посредством апикальной клетки. От них радиально расходятся многочисленные вильчато-разветвлен- ные ветви ограниченного роста, склеенные плотной слизью (рис. 1.35). Немалион служит типичным примером красной водоросли с многоосевым строением слоевища.
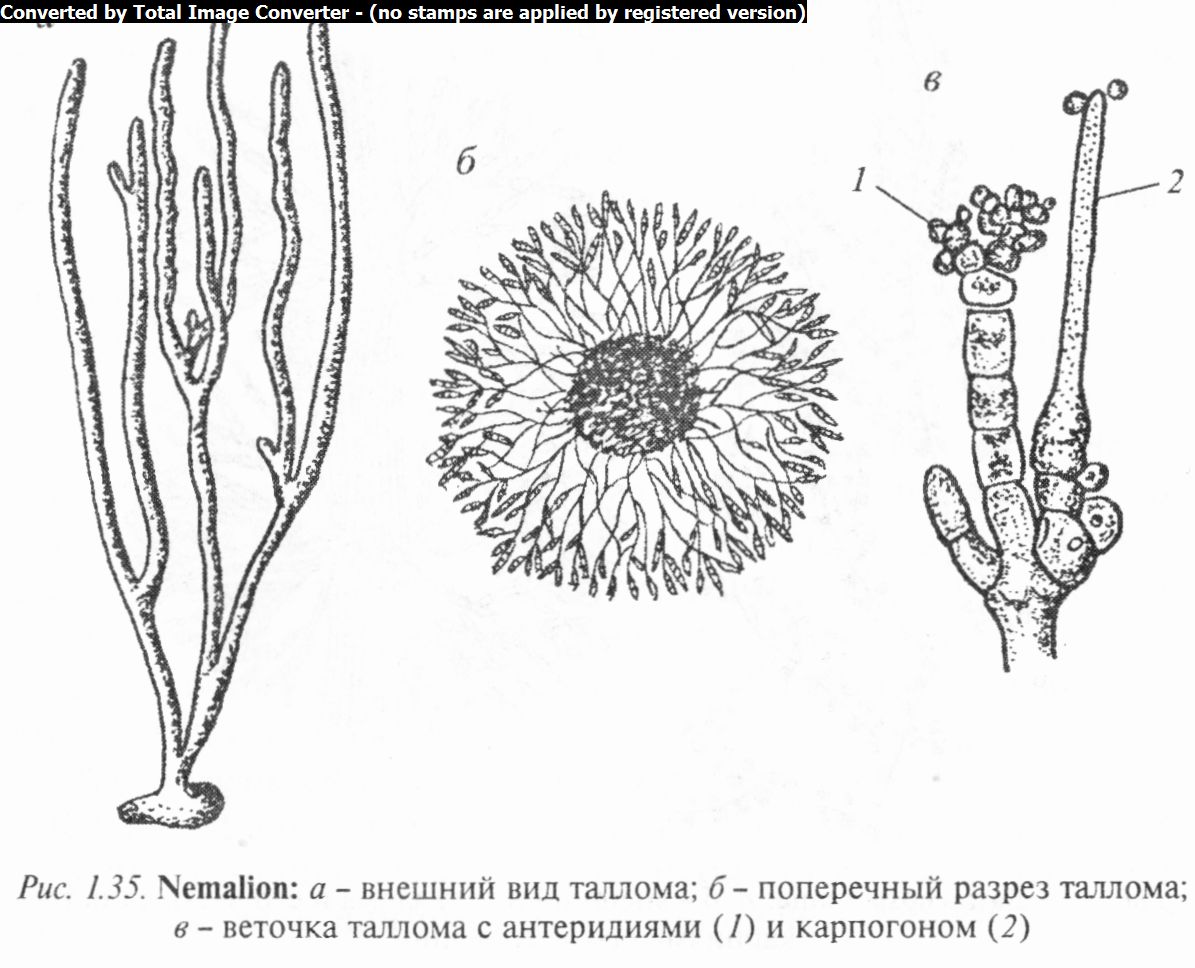
Органы полового размножения развиваются на ассимиляторах. Сперматангии гроздевидно расположены на конечных разветвлениях ассимиляторов. Карпогоны формируются обычно неподалеку от сперматангиев на карпогонных веточках. У одного из видов немалиона (N. vermiculare) наблюдалась гетероморфная смена поколений: макроскопического гаметофита и микроскопического нитевидного спорофита (тетраспорофита).
Порядок Церамиальные (Ceramiales). Наиболее высокоорганизованный и богатый видами порядок. Характеризуется тем, что ауксилярные клетки дифференцируются только после оплодотворения и в непосредственной близости от карпогона, т.е. имеется прокарпий. Цикл развития, как правило, трехфазный — со сменой гаметофита, карпоспорофита и тетраспорофита. Если репродуктивная система церамиальных построена по единому плану, то их вегетативная организация отличается большим разнообразием. На этом и основана классификация порядка. Все церамиальные построены по одноосевому плану.
Род каллитамнион (Callithamnion) включает виды, у которых таллом имеет форму разветвленного кустика, сложенного из одного ряда многоядерных клеток, различных по размеру у его основания и на вершине. Стенки клеток сильно ослизнены. На поперечных перегородках отчетливо видны поры, характерные для таллома красных водорослей (рис. 1.36, а).
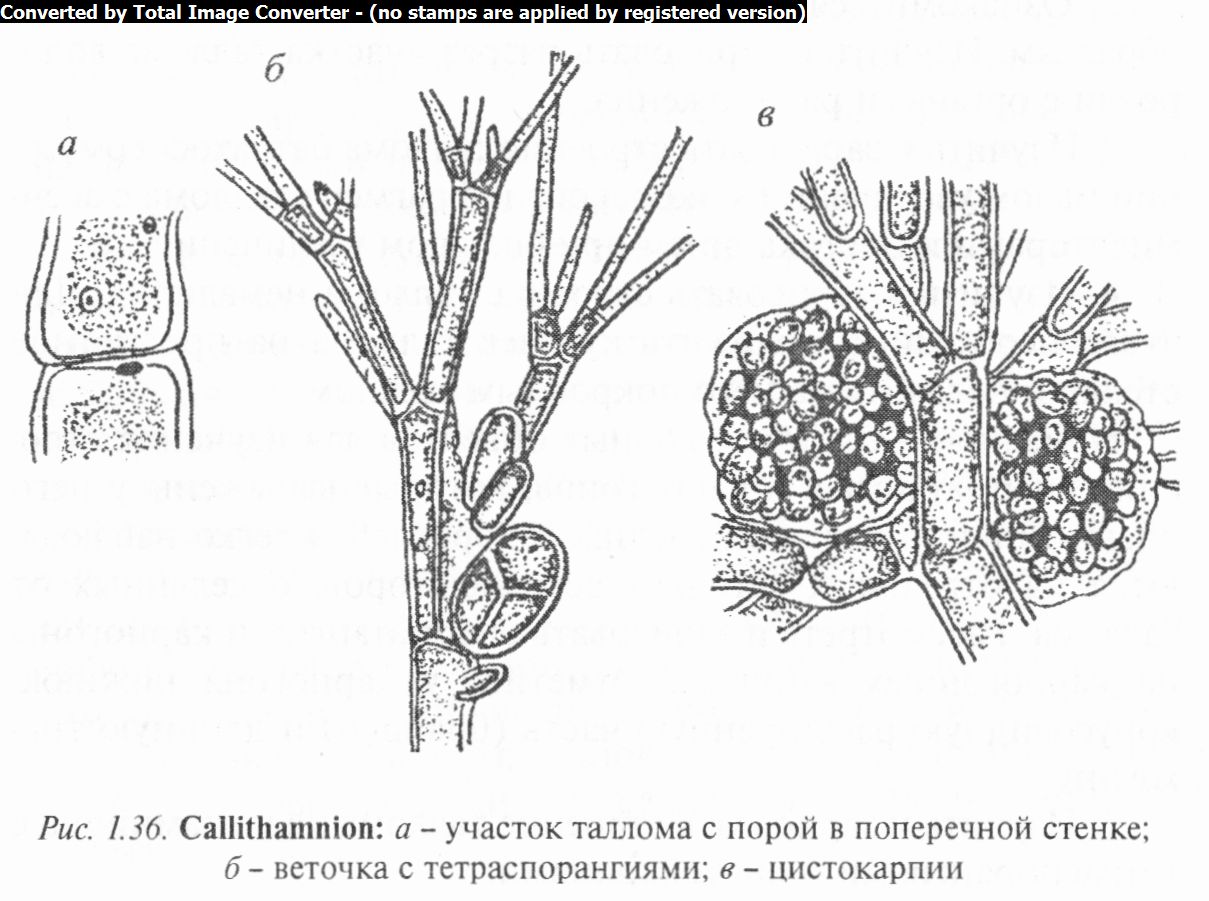
Некоторые виды имеют кору, которая состоит из ризо- идных нитей, спускающихся по стенкам основных ветвей слоевища.
Тетраспорангии образуются на верхних веточках в виде боковых овальных клеток, покрытых ослизненной оболочкой и сидящих на одноклеточных ножках. Споры располагаются по углам тетраэдра (рис. 1.36, 6).
Цистокарпии формируются на других экземплярах. Предварительно здесь возникают сперматангии и карпогоны. После оплодотворения из карпогона развиваются два цисто- карпия, расположенные супротивно по бокам нитевидного таллома. Кучки карпоспор часто покрыты общей слизистой оберткой (рис. 1.36, в).
Известно более 50 видов, распространенных в различных морях. Несколько видов обитает в Черном море, среди них наиболее часто встречается К. щитковидный (С. corymbosum).
Задания
1. По гербарным образцам ознакомиться с разнообразием талломов багрянок, обратив внимание на их размеры, форму, окраску и др.
2. Ознакомиться с общим видом порфиры по гербарным образцам. Изучить и зарисовать разрез участка таллома водоросли с органами размножения.
3. Изучить и зарисовать строение таллома батрахоспермума при малом увеличении микроскопа и фрагмент таллома с ассимиляторами и цистокарпием при большом увеличении.
4. Изучить и зарисовать строение таллома немалиона. Для этого необходимо положить кусочек таллома на предметное стекло и надавить на него покровным стеклом.
Немалион — один из удобных объектов для изучения строения сперматангиев и карпогонов, которые выражены у него очень типично для этой группы водорослей и легко наблюдаются на отдельных веточках ассимиляторов, отделенных от таллома. Рассмотреть и зарисовать сперматангии и карпогоны на карпогонных веточках. Отметить у карпогона нижнюю конусовидную расширенную часть (брюшко) и длинную три- хогину.
5. Изучить и зарисовать строение таллома каллитамниона с тетраспорангиями и цистокарпиями.
Вопросы и задания для самоконтроля
1. Каковы особенности строения клетки и таллома красных водорослей?
Дата добавления: 2021-03-18; просмотров: 909;
Поиск по сайту
Узнать еще
- II. Бесполые способы размножения.
- III. Суммарные допуски формы расположения
- V. Сборка и нагружение формы.
- V.VII. Зависимость ширины и формы выхода слоя на поверхности от его истинной мощности, угла падения и формы рельефа
- А - с прямолинейной спинкой; б - с криволинейной спинкой; в - с канавкой удлиненной формы
- А) Контроль отклонений от правильной цилиндрической формы.
- А. Локализованные формы
- Автоматический контроль точности размеров и формы деталей. Разновидности контроля. Использование информации, полученной при контроле,

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
