Порядок Хетофоральные
(Chaetophorales). Включает раз- нонитчатые ветвящиеся формы. Подавляющее большинство водорослей этого порядка имеет на слоевище волоски или щетинки. Бесполое размножение происходит при помощи четырех- жгутиковых зоо- и апланоспор.
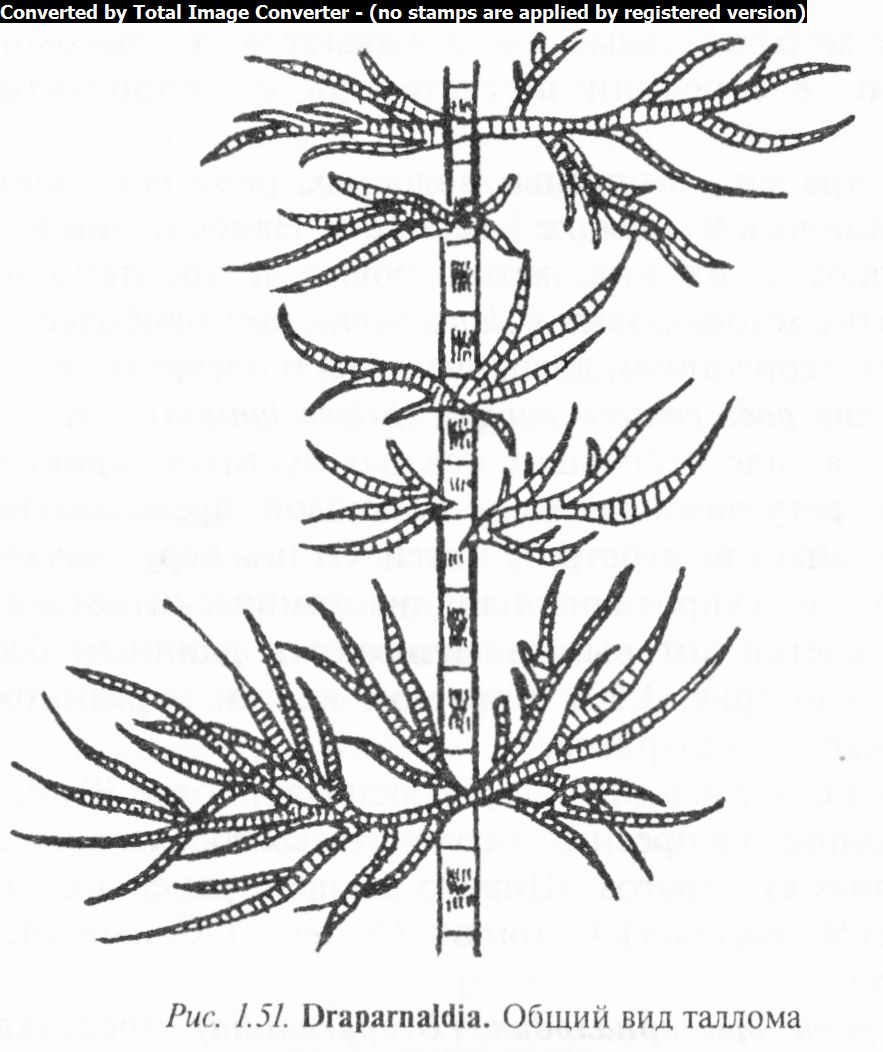
волосками. Прикрепляется драпарнальдия ризоидом (рис. 1.51). Половой процесс изогамный или гетерогамный, бесполое размножение осуществляется четырехжгутиковыми зооспорами.
Известно 19 видов драпарнальдии, распространенных в прибрежной полосе озер, проточных холодных вод и растущих на разных субстратах. В Беларуси 2 вида. Широко распространены Д. скрученная (D. glomerata), Д. перистая (D. plumosa), Д. простая (D. simplex) и др.
Водоросли рода плеврококк (Pleurococcus) встречаются в виде зеленого порошистого налета на коре деревьев, на стенах, камнях и старых заборах. Клетки плеврококка округлые, одиночные или сросшиеся вместе по 3 — 4 и более, часто образуют характерные пакетики. Они одеты довольно толстой целлюлозной оболочкой с одним хроматофором, как правило, без пиреноида (рис. 1.52).
Размножение только вегетативное. Материнские оболочки после деления срастаются с дочерними. Часто в результате деления в одном направлении образуются нити, поэтому многие систематики считают плеврококк многоклеточной водорослью, имеющей редуцированный гетеротрихальный таллом.
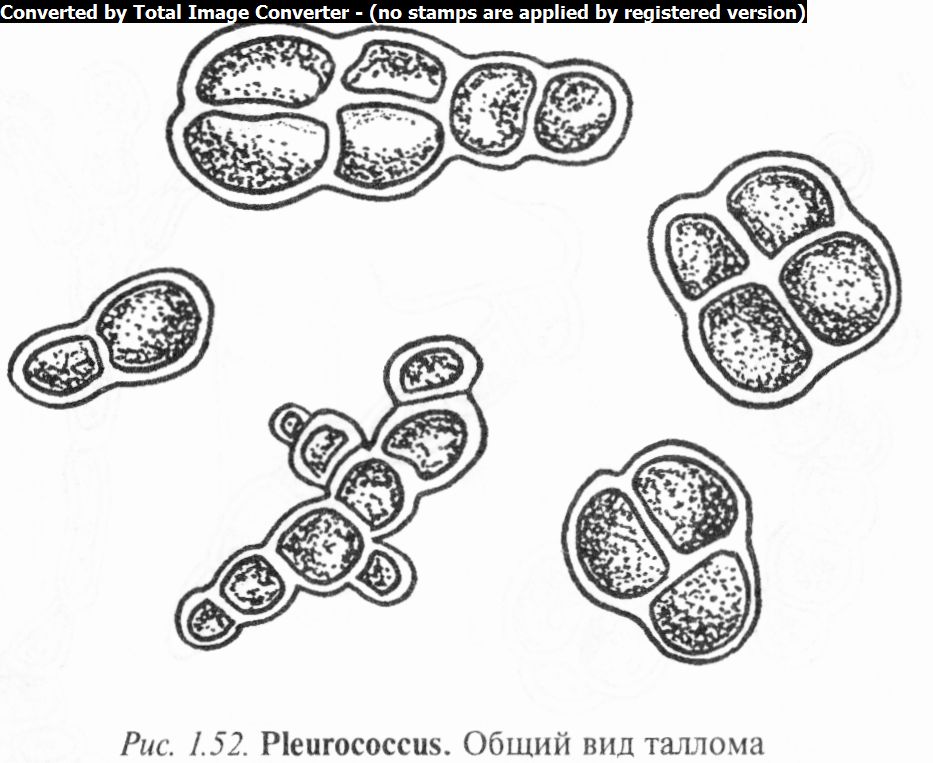
Известно 5 видов. В Беларуси чаще других встречается П. обыкновенный (P. vulgaris).
Род трентеполия (Trentepohlia) одноименного семейства включает аэрофильные водоросли, обитающие на коре деревьев, камнях, деревянных зданиях, образуя на них налет кирпич- но-красного или желтого цвета (за счет гематохрома, растворенного в каплях масла). Гетеротрихальный таллом состоит из стелющихся по субстрату и вертикальных нитей. Первые — короткие, разветвленные, без ризоидов, нередко распадающиеся на отдельные клетки, которые разносятся токами воздуха или воды, обеспечивая тем самым вегетативное размножение и расселение. Клетки их овальные, одеты толстыми, часто слоистыми оболочками. Хроматофоры в виде дисков, без пиреноидов. На стелющихся нитях образуются шаровидные или эллипсовидные гаметангии (рис. 1.53).
Восходящие нити образованы более вытянутыми цилиндрическими клетками. На концах этих нитей образуются овальные или шаровидные зооспорангии. Они легко отделяются и переносятся ветром или водой. Размножение бесполое, двух-, четырехжгутиковыми зооспорами, половое — двухжгути- ковыми изогаметами, которые развиваются в сидячих гаметан- гиях. Гаметы копулируют не всегда. Они могут прорастать в нити, как зооспоры, либо превращаться в апланоспоры. Вегетативное размножение осуществляется участками нитей.
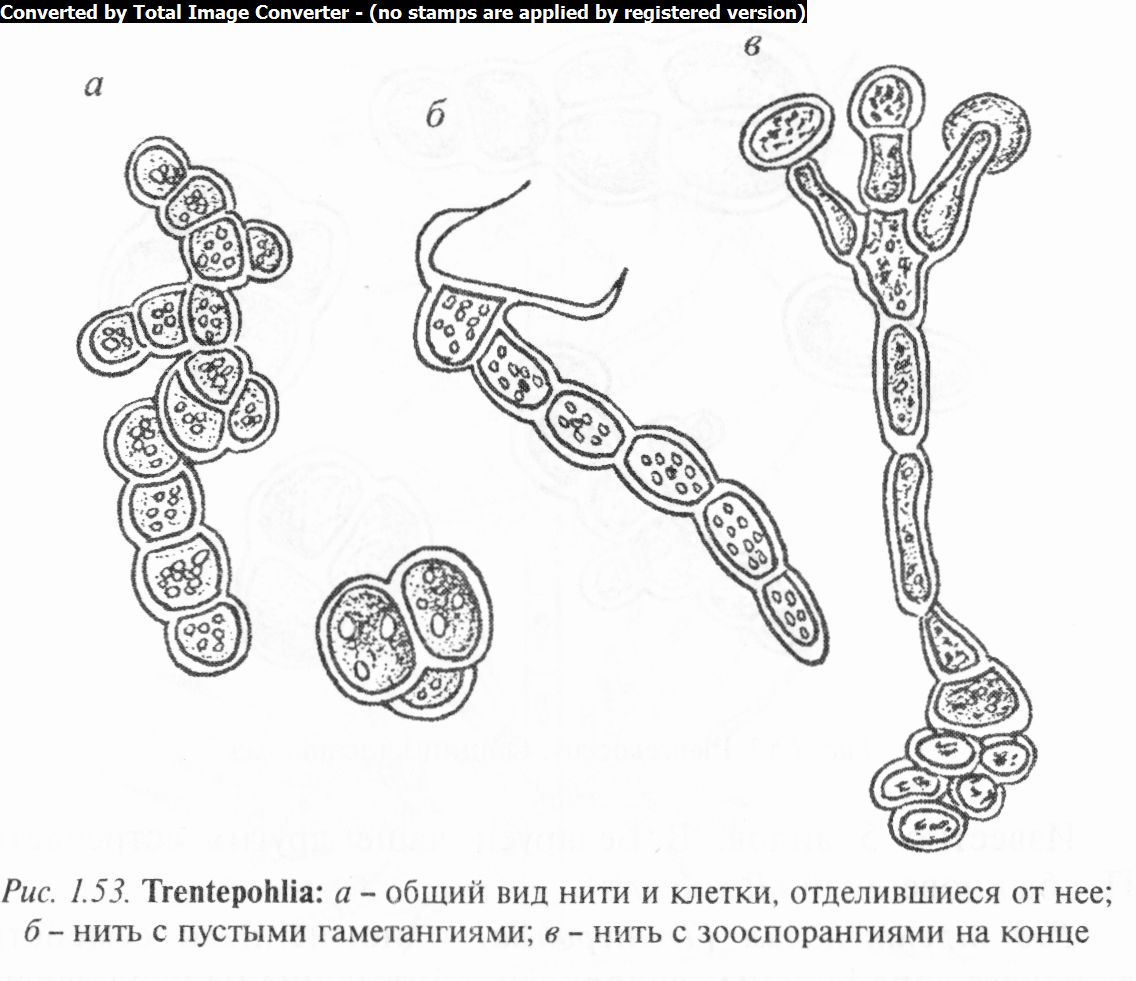
Известно около 60 видов трентеполии, в Беларуси их 4. Широко распространены Т. затененная (Т. umbrina), Т. золотистая (Т. аигеа), Т. смолистая (Т. piceana) и др.
Порядок Эдогониальные (Oeclogoniales). Он объединяет нитчатые, одноядерные, преимущественно неветвящиеся прикрепленные формы, с особыми «колпачками», которые образуются на оболочках клеток при их делении. Зооспоры и сперматозоиды многожгутиковые, жгутики располагаются венцом. Половой процесс оогамный. Порядок включает одно семейство эдогониевых (Oedogoniaceae), представленное тремя родами.
Род эдогониум (Oeilogonium) характеризуется тем, что его слоевище построено по типу длинных неразветвленных нитей, прикрепляющихся в молодом состоянии к субстрату с помощью базальной клетки; зрелые нити обычно свободноплавающие. Нити состоят из цилиндрических клеток с гладкими оболочками. Верхушечные клетки часто заостряются и заканчиваются щетинкой. Клетки содержат крупное ядро и пристенный сетчатый хроматофор со многими пиреноидами.
Характерным признаком, по которому эдогониум можно легко отличить от других нитчатых, служат «колпачки», форми
рующиеся на делящихся клетках (рис. 1.54). Особенно резко они заметны на пустых, мертвых клетках или над оогониями в виде поперечных штрихов на оболочке у верхнего конца клет-
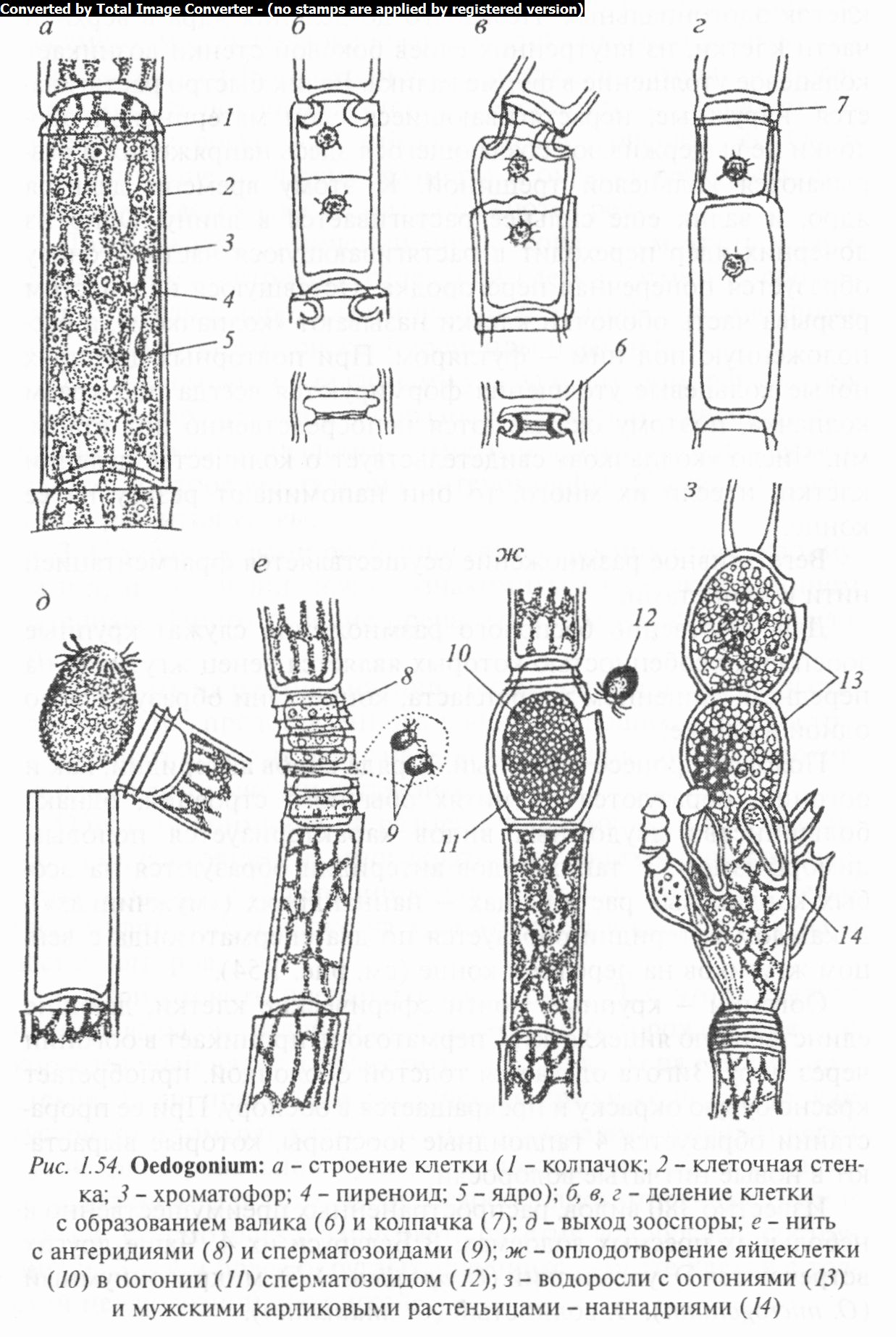
ки. По бокам в этих местах оболочка имеет как бы зазубренный край.
Возникают «колпачки» в результате своеобразного деления клеток эдогониальных. Незадолго до деления ядра в верхней части клетки, из внутренних слоев боковой стенки возникает кольцевое утолщение в форме валика. Валик быстро растягивается, наружные, нерастягивающиеся слои материнской оболочки не выдерживают создающегося здесь напряжения и разрываются кольцевой трещиной. К этому времени делится ядро, и валик еще сильнее растягивается в длину. Одно из дочерних ядер переходит в растягивающуюся часть, а снизу образуется поперечная перегородка. Оставшуюся над местом разрыва часть оболочки клетки называют «колпачком», а расположенную под ним — футляром. При повторных делениях новые кольцевые утолщения формируются всегда под краем колпачка, поэтому оказываются непосредственно под старыми. Число «колпачков» свидетельствует о количестве делений клетки, и если их много, то они напоминают резьбу на ее конце.
Вегетативное размножение осуществляется фрагментацией нити и акинетами.
Для собственно бесполого размножения служат крупные зооспоры, особенностью которых является венец жгутиков на переднем, лишенном хлоропласта, конце. Они образуются по одной в клетке.
Половой процесс оогамный. У ряда видов антеридии, как и оогонии, образуются на нитях обычного строения, однако большинство двудомных видов характеризуется половым диморфизмом. У таких видов антеридии образуются на особых карликовых растеньицах — наннандриях («мужчинках»), В каждом антеридии образуется по два сперматозоида с венцом жгутиков на переднем конце (см. рис. 1.54).
Оогонии — крупные, почти сферические клетки, дающие единственную яйцеклетку. Сперматозоид проникает в оогонии через пору. Зигота одевается толстой оболочкой, приобретает красно-бурую окраску и превращается в ооспору. При ее прорастании образуется 4 гаплоидные зооспоры, которые вырастают в новые нитчатые водоросли.
Известно 380 видов, распространенных преимущественно в небольших пресных водоемах. В Беларуси их 4. Чаще других встречаются Э. упсальский (О. upsaliense), Э. микрогониумный (О. microgonium), Э. волнистый (О. undulatum).
Задания
1. Приготовить препарат улотрикса. Для этого следует взять небольшое количество нитей и поместить в каплю воды на предметном стекле. Покачиванием стекла можно добиться их равномерного распределения, не повредив материала, после чего накрыть покровным стеклом. Рассмотреть при малом увеличении микроскопа общий вид нити и зарисовать ее. Обратить внимание на форму, размеры и окраску верхушечной и базальной клеток (последние попадаются редко).
1. При большом увеличении микроскопа рассмотреть и зарисовать форму и строение отдельной клетки. Отметить оболочку, цитоплазму, ядро, пластинчатый хроматофор в виде широкого незамкнутого пояска, пиреноиды и вакуоль.
2. Ознакомиться с талломами ульвы и энтероморфы на гербарных и фиксированных образцах. Изучить и зарисовать внутреннее строение талломов, отметить двухслойность ульвы и трубчатое строение таллома энтероморфы. Составить схему цикла развития ульвы.
3. С характерными особенностями строения стигеоклониу- ма и драпарнальдии можно ознакомиться на фиксированном материале. Приготовление препарата такое же, как и для улотрикса.
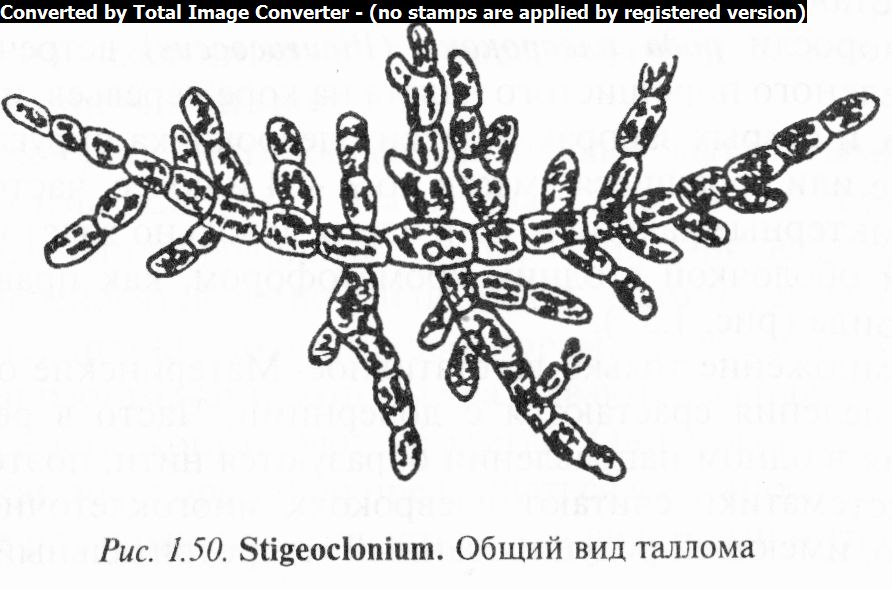
3. Отметить на рисунке наличие ризоидов или подошвы у сти- геоклониума, представленного в виде стелющихся по субстрату нитей, от которых берут начало восходящие нити, супротивно или дихотомически ветвящиеся.
3. Обратить внимание на размеры и форму кустиков драпарнальдии, состоящих из многократно ветвящихся одноядерных нитей с ясной дифференцировкой на ствол и боковые ветви (ассимиляторы), а также на хроматофоры в клетках ствола и ассимиляторов.
4. Приготовить препарат плеврококка. Для этого следует препаровальной иглой или скальпелем осторожно снять с коры зеленый налет и поместить в каплю воды на предметном стекле. Если при малом увеличении препарат выглядит как густой конгломерат клеток, то нужно надавить пальцем через кусок фильтровальной бумаги на покровное стекло и растереть материал для того, чтобы освободились отдельные нити и клетки. Рассмотреть и зарисовать пакетик водорослей, состоящих из 3 — 4 и более округлых, сросшихся вместе клеток с толстой целлюлозной оболочкой.
5. Приготовить препарат трентеполии таким же способом, как и плеврококка.
6. Изучить и зарисовать гетеротрихальное строение таллома трентеполии. Отметить различие в строении клеток стелющихся по субстрату и восходящих нитей. На первых из них найти и зарисовать шаро- или эллипсовидные гаметангии, на вторых — овальные или шаровидные зооспорангии, расположенные на особых субспорангиальных клетках.
7. Рассмотреть под микроскопом и зарисовать нить эдого- ниума. Отметить наличие «колпачков», которые следует искать при большом увеличении. Особенно резко они видны на пустых, мертвых клетках или над оогониями в виде поперечных штрихов на оболочке у верхнего конца клетки. Обратить внимание на расположение, размеры, форму и строение органов полового размножения — оогония и антеридия.
Вопросы и задания для самоконтроля
1. Каковы особенности строения и жизненного цикла улотрикса?
2. Какие водоросли порядка улотриксовых отклоняются от характерного для зеленых водорослей жизненного цикла?
3. Охарактеризуйте водоросли, имеющие гетеротрихальное строение таллома.
4. Каковы пути эволюции в развитии гетеротрихального таллома? Дайте примеры существующих водорослей.
5. В чем особенности деления клетки и размножения эдогониальных?
6. В чем сходство и различие улотрихальных и протококальных водорослей?
7. Каковы особенности строения и размножения хетофоральных водорослей?
Класс Сифонофициевые (Siphonophyceae)
Класс включает водоросли, у которых таллом имеет сифоновую и сифонокладальную структуру. Их слоевище представляет собой гигантскую одно- или многоядерную клетку или сегментировано своеобразными перегородками на отдельные участки с многочисленными ядрами. Несмотря на отсутствие клеточных перегородок, они, по сути дела, не являются одноклеточными организмами. Их слоевище скорее напоминает комплекс не вполне разделившихся клеток. Другими словами, увеличение количества ядер можно рассматривать как резуль тат незаконченного процесса клеточного деления, когда ядро делится, а цитоплазма остается неразделенной.
Оболочка клеток целлюлозная, каллозная или пектиновая, у некоторых форм пропитана известью. В постенном слое цитоплазмы находятся многочисленные дисковидные хроматофоры обычно без пиреноидов, а под ними — ядра. Если хроматофор один, он имеет сетчатое строение. В хроматофорах кроме обычных пигментов, свойственных зеленым водорослям, имеются специфические — сифонеин и сифоноксантин из группы каротиноидов.
Сифонофициевым водорослям свойственны все типы размножения: вегетативное (выводковые почки, акинеты), собственно бесполое (зоо- и апланоспоры) и половое (изо-, гетеро- и оогамия). Они дипло- или диплогаплобионты с изоморфной, реже гетероморфной сменой форм развития.
Класс делится на 3 порядка (Сифональные, Дазикладальные и Сифонокладальные), каждый из которых представляет собой самостоятельную эволюционную линию.
Сифонофициевые обитают главным образом в тропических морях.
Порядок Сифональные (Siphonales), или Бриопсидальные(Biyopsidules). Слоевище водорослей лишено перегородок, только в основании ветвей и изредка гаметангиев возникают своеобразные пробки. В пристенном слое цитоплазмы содержатся многочисленные ядра и хроматофоры с пиреноидами или без них.
Размножение осуществляется преимущественно половым путем. Зооспоры известны лишь у немногих видов. Некоторые могут размножаться вегетативно отрывающимися или распадающимися при разрастании участками тела, легко регенерирующими: оболочка в местах отрыва очень быстро восстанавливается. В порядке 3 семейства: бриопсидовые, кодиевые и каулерповые.
Род бриопсис (Bryopsis) одноименного семейства объединяет виды, у которых слоевище состоит из коротких малоразветв- ленных ризомов и системы довольно крупных вертикальных побегов, имеющих главную ось с веточками первого и второго порядков. У одних видов ветви располагаются супротивно и в одной плоскости (таллом напоминает перо птицы), у других — вокруг главной оси (схожи с метелкой злаков) (рис. 1.55). Вегетативное размножение происходит путем отделения «перышек», которые закупориваются в точке отрыва и затем выра-
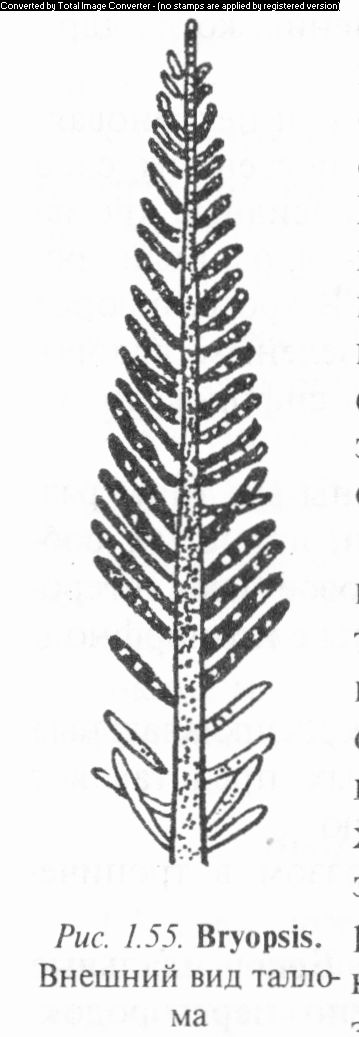
стают в новые организмы. Кроме того, участки протопласта, вытекающие из таллома при его повреждениях, округляются, формируют оболочку и развиваются в новую водоросль. Некоторые представители образуют выводковые почки. Собственно бесполое размноже-
обнаружено лишь у тех видов, у которых 1 гетероморфная смена форм развития. Осуществляется оно крупными шаровидными зооспорами с несколькими ядрами, хромато- форами и венцом жгутиков.
Половой процесс — изо- или гетерогам- ный, как и у других сифонофициевых. Гаметы образуются в конечных веточках, которые, перед тем как стать гаметангиями, не меняя формы, отделяются перегородкой от слоевища. Вышедшие из гаметангиев мужские и женские гаметы попарно копулируют, и образовавшаяся зигота немедленно начинает про- Рис. 1.55. Bryopsis. растать в первичную нить (протонему), из Внешний вид талло-которой развиваются гаметофиты или ма зооспоры. В последнем случае протонема
функционирует как спорофит. Таким образом, у одних видов бриопсиса в цикле развития доминирует гаплоидная фаза (диплоидна лишь зигота), у других же происходит гетероморфная смена поколений.
Известный вид Б. перистый (В. plumosa) обитает в Черном, Азовском и других морях, поселяется на скалах, камнях, раковинах и т.д.
Род кодиум (Codium) (рис. 1.56) одноименного семейства содержит 50 видов, которые характеризуются тем, что талломы, имеющие типичное сифоновое строение, плотно сплетаются, образуя компактные псевдопаренхиматозные слоевища.
В талломе имеются две зоны. Внутренняя зона образована пучком тонких бесцветных нитей, от которых к периферической части отходят кортикальные пузыри, или утрикулы, образующие поверхностный слой таллома. Утрикулы являются своеобразными ассимиляторами. Оболочки клеток каллозопекти- новые с небольшой примесью целлюлозы. Размножается вегетативно — отрезками слоевища и специальными пузырьками, развивающимися у основания таллома (см. рис. 1.56).
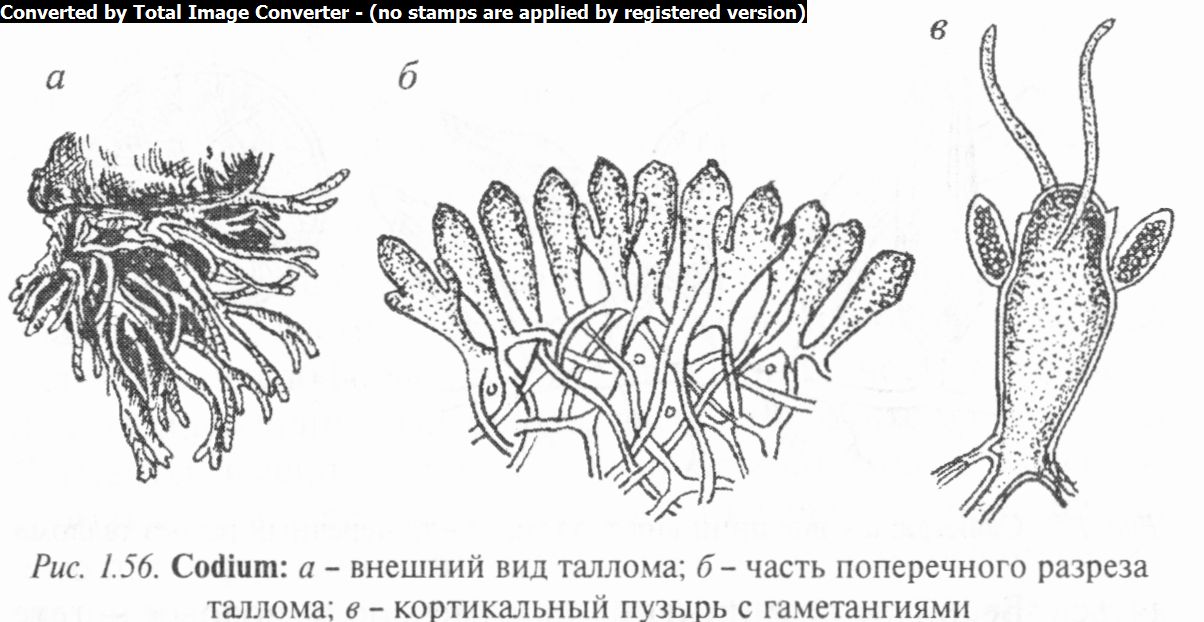
Половой процесс — гетерогамный. Гаметангии формируются по бокам утрикул. Мужские и женские гаметангии можно различить еще до того, как образуются гаметы, благодаря золо- тисто-желтой окраске первых и темно-зеленой окраске вторых. Зигота непосредственно прорастает в новую водоросль.
Кодиевые являются более высокоорганизованными представителями сифонофициевых, что проявляется в псевдопаренхиматозном строении таллома, дифференцировке гаме- тангиев и полной утрате бесполого размножения.
Виды кодиума встречаются в тропических морях, реже в более холодных. Поселяются на различных твердых грунтах.
Род каулерпа (Caulerpa) одноименного семейства объединяет виды, у которых слоевище расчленяется на стелющиеся трубковидные ризомы с ризоидами и вертикальные ассимиляционные побеги. В талломе развивается своеобразный внутренний скелет в виде радиальных или перпендикулярных тяжей, которые увеличивают механическую прочность клетки.
Особенностью каулерпы является наличие в составе оболочки каллозы (вместо целлюлозы), пектиновых и других веществ, а также лейкопластов, участвующих в накоплении крахмала.
В роде 60 видов, обитающих главным образом в тропических и субтропических морях.
Каулерпа прорастающая (С. prolifera) — типичный представитель рода (рис. 1.57). Ризомы ее могут достигать в длину 1 м, а вертикальные листовидные побеги не превышают 30 см. Размножается каулерпа за счет отрастания новых побегов, а также путем отделения частей слоевища, способных вновь прикреп
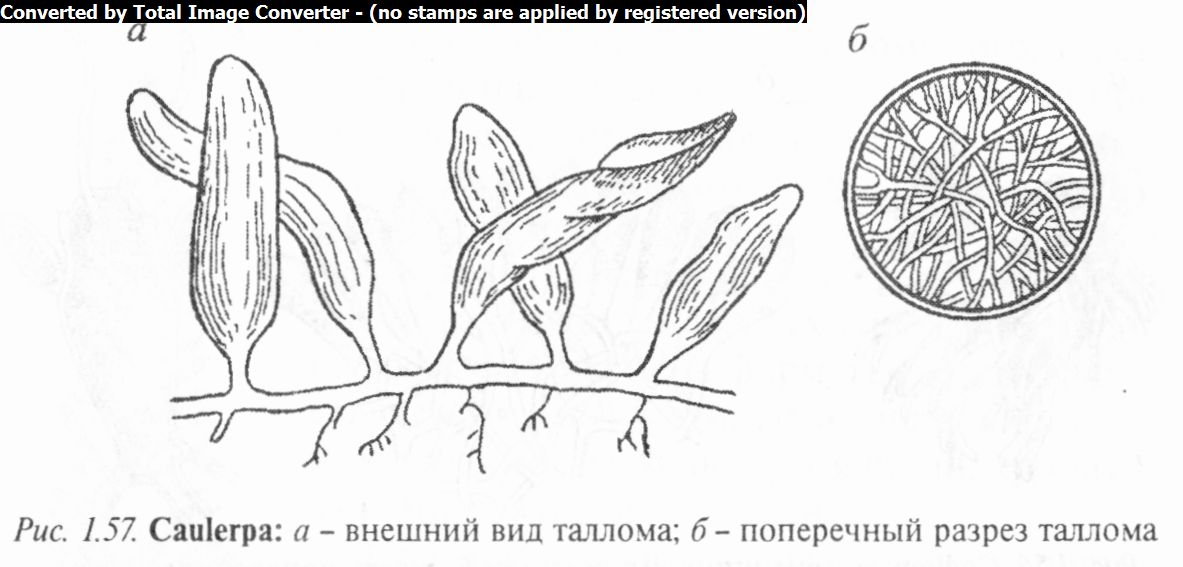
ляться. Бесполое размножение не отмечено, а половое - гете- рогамное. Гаметы образуются в любой ассимиляционной части таллома. При этом часть многоядерной цитоплазмы сгущается, становится темно-зеленой. Ядра делятся редукционно, в результате чего образуются одноядерные двухжгутиковые гаметы, выходящие из таллома через длинные выросты, — папиллы. Затем гаметы сливаются, а образовавшаяся зигота вскоре прорастает в новую особь. Такое размножение называется голокарпией. В цикле развития каулерпы доминирует диплоидная фаза. Гаплоидны лишь гаметы.
Обитает каулерпа на скалах, камнях, в песчаном грунте Средиземного, Черного и других морей.
Порядок Сифонокладальные (Sip ho по с la da les). Порядок объединяет водоросли, имеющие сегментированное слоевище. Сегменты образуются за счет сегрегативного деления цитоплазмы на многоядерные участки различной формы и размеров. Каждый из этих участков вырабатывает собственную мембрану. Разрастаясь, они смыкаются, создавая впечатление многоклеточности. Таким образом, деление цитоплазмы и образование сегментов у них не связано с делением ядер. Все сифонокладальные на начальных фазах развития проходят стадию первичного пузыря, что свидетельствует об их родстве с сифональными. Сегментация наступает позже. Строение сегмента сифонокладальных сходно со строением клетки сифо- нальных водорослей.
Размножаются сифонокладальные половым и бесполым путем. Имеется изо- и гетероморфная смена форм развития.
Распространены главным образом в тропических морях и лишь некоторые в пресных водоемах.
Порядок включает несколько семейств, различающихся строением слоевища. Наиболее высокоорганизованным и широко распространенным является семейство кладофоровых (Cladophoraceae) с типичным родом кладофора.
Род кладофора (Claclophora) (рис. 1.58) объединяет виды, у которых слоевище представляет собой разветвленные сифонокладальные талломы кустистой или шаровидной формы. Каждая ветка кладофоры состоит из цилиндрических, бочонко- или эллипсовидных сегментов, располагающихся в один ряд. Сегменты имеют целлюлозные оболочки, многочисленные хроматофоры, в результате смыкания которых образуются сет- чато продырявленные постенные пластинки с многочисленными пиреноидами. Цитоплазма содержит по нескольку ядер. В центре сегмента находится крупная вакуоль.
Вегетативное размножение осуществляется частями таллома или акинетами.
При собственно бесполом размножении кладофоры в конечных клетках ее ветвей, выделяющихся своим густо-зеле- ным цветом, образуется масса мелких четырехжгутиковых
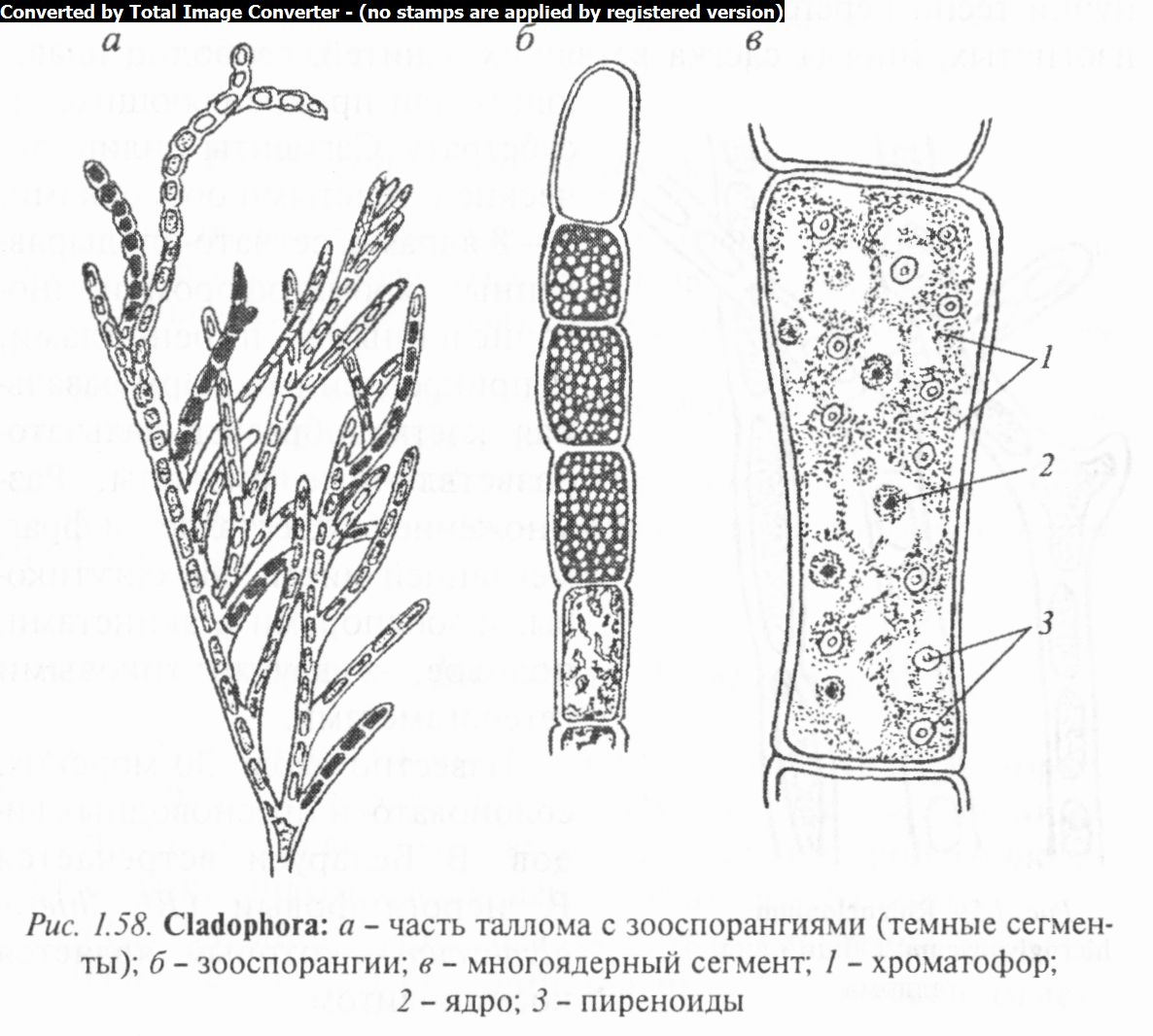
зооспор. Половое размножение изогамное, при помощи двух- жгутиковых гамет, которые формируются в любом сегменте водоросли. У пресноводных видов доминирует диплоидная фаза, зооспоры диплоидны. В цикле развития этих видов осуществляется лишь смена ядерных фаз. Из зиготы морских видов вырастает спорофит, внешне сходный с гаметофитом, но продуцирующий четырехжгутиковые зооспоры. В цикле их развития происходит изоморфная смена поколений. При прорастании зооспоры и зиготы вначале формируется сифональ- ная структура булавовидного слоевища и происходит диффе- ренцировка его на базальную и апикальную части. Позже апикальная часть сегментируется. Таким образом, онтогенез кла- дофоровых свидетельствует об их родстве с сифональными.
Известно около 150 видов, обитающих в морях и пресноводных водоемах, в Беларуси их 4. Широко распространены К. речная (С. rivularis), К. скрученная (С. glomerata) и др. Кладофора эгагропильная (С. aegagropila) занесена в Красную книгу Республики Беларусь.
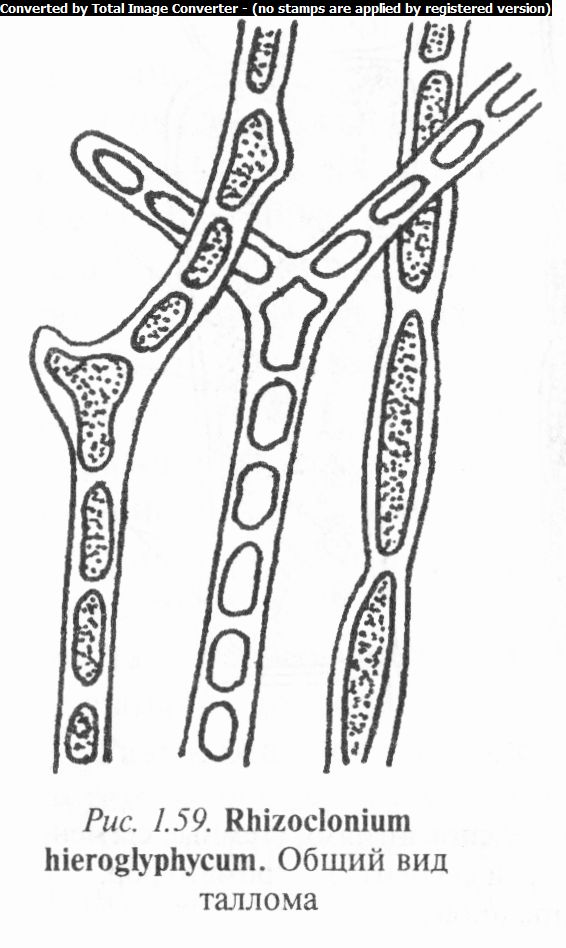
Виды рода ризоклониум (Rhizoclonium) (рис. 1.59) образуют пучки тесно переплетающихся длинных, тонких, прямых или изогнутых, иногда слегка ветвящихся нитей, свободно плавающих или прикрепляющихся к
субстрату. Сегменты цилиндрические с толстыми оболочками, 1 — 8 ядрами, сетчато-продырявленным хроматофором и многочисленными пиреноидами. У прикрепленных форм базальная клетка образует пальчато- разветвленные ризоиды. Размножение осуществляется фрагментацией нитей, двухжгутиковыми зооспорами и акинетами, половое — двухжгутиковыми гетерогаметами.
Известно около 30 морских, солоновато- и пресноводных видов. В Беларуси встречается Р. иероглифовый (Rh. hiero- glyphycum), который является космополитом.
Задания
1. Ознакомиться с внешним видом кладофоры. Обратить внимание на крупные войлочные дерновинки темно-зеленого цвета, жесткость нити водоросли из-за отсутствия слизистого чехла. Приготовить препарат, рассмотреть при малом увеличении микроскопа и зарисовать часть ветвящегося таллома. Отметить дифференцировку тела на главную ось и боковые ветви, на сегменты с зооспорангиями и гаметангиями.
2. Рассмотреть при большом увеличении микроскопа и зарисовать отдельный сегмент кладофоры. Отметить толстую целлюлозную оболочку, цитоплазму, сетчато-продырявленный хроматофор с многочисленными пиреноидами, крупные ядра. Для того чтобы лучше рассмотреть ядра, рекомендуется очень выразительная быстрая окраска ацетокармином. После этого препарат желательно рассматривать под микроскопом с иммерсионным объективом.
3. С представителями рода бриопсис, кодиум, каулерпа, ризоклониум и другими сифональными рекомендуется ознакомиться на фиксированном материале и по гербарным образцам. Обратить внимание на размеры и форму тела, окраску и характер ветвления таллома этой группы водорослей.
Вопросы и задания для самоконтроля
1. Охарактеризовать особенности строения, размножения и цикла развития сифональных водорослей на примере бриопсиса и каулерпы.
2. Чем отличается сифональная структура таллома от сифонокладаль- ной?
3. Каковы особенности размножения и смены ядерных фаз у представителей порядка Сифонокладальные?
4. Какие особенности сифональных позволяют использовать их в качестве модельных объектов при изучении ядерно-цитоплазматических взаимоотношений?
Класс Конъюгатофициевые (Conjugatophyceae)
Класс объединяет водоросли, характерной чертой которых являются полная потеря подвижных жгутиковых стадий и половой процесс в виде конъюгации, смысл которой заключается в слиянии протопластов двух вегетативных клеток.
Это одно- и многоклеточные нитчатые организмы, изредка колониальные. Таллом большинства их видов имеет симмет ричное строение с перетяжкой посередине или без нее. У нитчатых форм клетки не перешнурованы. Ободочки клеток целлюлозные, ослизняющиеся. Протопласты содержат по одному гаплоидному ядру, крупные осевые или постенные хроматофоры, имеющие пластинчатую, лентовидную, звездчатую, спиралевидную форму, пиреноиды.
Размножаются конъюгаты бесполым и половым путями.
Бесполое размножение осуществляется за счет деления клеток и распада нитей на отдельные участки и даже на клетки, способные регенерировать всю нить. Кроме того, изредка могут формироваться апланоспоры, партеноспоры, а также акинеты, служащие для переживания неблагоприятных условий и размножения.
Половой процесс — конъюгация - представляет собой слияние амебоидных протопластов вегетативных клеток. Если скорость их перетекания одинакова, то слияние происходит в копуляционном канале. В этом случае процесс условно называют изогамным. Если же протопласты двух клеток перетекают с разной скоростью, слияние происходит в одной из них и тогда воспринимающую клетку называют женской, а клетку с более подвижным протопластом — мужской. Половой процесс в этом случае носит название физиологической гетерогамии. После периода покоя зиготы прорастают 1 — 4 жизнеспособными проростками, что сопровождается редукцией числа хромосом. В цикле развития доминирует гаплоидная фаза, диплоидны лишь зиготы.
Известно около 4700 видов, распространенных на всех континентах.
Конъюгаты — космополиты. Они приурочены преимущественно к пресным водоемам. Имеются также наземные виды, встречающиеся на сырой земле, скалах, мхах, ледниках высокогорий.
В классе 4 порядка: Мезотениальные, Зигнематальные, Десми- диальные и Гонатозигальные. В основе такой классификации лежат строение клеток и количество жизнеспособных проростков.
Порядок Мезотениальные (Mesotaeniales). Представители этого порядка одноклеточные, коккоидные, со сплошными однослойными гладкими оболочками без пор, покрытые слизистой оберткой или чехлом. Особенно ослизнены неводные обитатели. Клетки могут быть цилиндрической, эллипсовидной или веретеновидной формы, без перетяжки. Хроматофоры имеют разную форму и размеры с пиреноидами. Ядро одно.
Размножаются мезотениальные бесполым (делением клетки надвое) и половым (конъюгация) путями. Зиготы после периода покоя прорастают 4 жизнеспособными проростками. По этой причине мезотениальные считают самыми примитивными среди конъюгат.
К мезотениальным относится 6 родов и 47 видов, около 20 отмечены в Беларуси.
Обитают мезотениальные преимущественно в торфяных водоемах. На севере некоторые из них вызывают красное «цветение» снега.
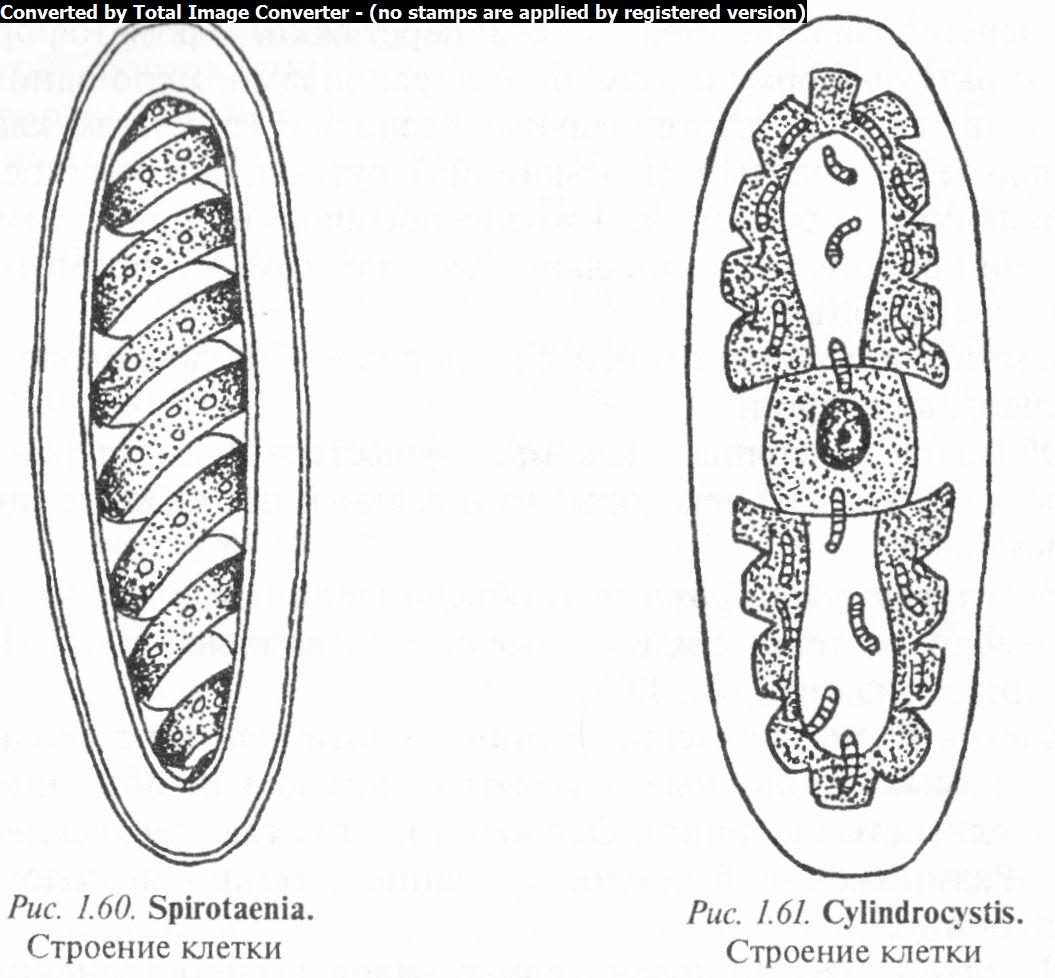
Род спиротения (Spirotaenia) объединяет виды, клетки кото- рызГймеют постенные или осевые спиралевидные хроматофоры с пиреноидами (рис. 1.60).
Клетки цилиндрические, эллипсовидные или веретеновид- ные, прямые, одиночные, покрытые слизью или объединенные в слизистые колонии. Оболочки клеток гладкие, бесцветные. Размножение бесполое (деление клетки) и половое (конъюгация).
Известно более 30 пресноводных видов, распространенных преимущественно в заболоченных водоемах, канавах, лужах. В Беларуси их 2 — С. сжатая (5. condensata) и С. темная (S. obscura).
Род цилиндроцистис (Cylindrocystis) (рис. 1.61) характеризуется двумя осевыми звездчатыми хроматофорами с пиреноидом в каждом из них. Клетки эллипсовидные или цилиндрические, с закругленными концами. Изредка окружены слизью. Оболочка гладкая, бесцветная, тонкая. Размножение бесполое (поперечное деление клеток) и половое (конъюгация).
Известно 5 видов цилиндроцистиса, распространенных преимущественно в водах с низкой кислотностью, во влажных местах и среди мхов. В Беларуси отмечены 2 вида — Ц. толстый (С. crassa), Ц. Бребиссона (С. brebissonii).
Порядок Зигнематальные (Zygnematales). Порядок включает нитчатые, неветвящиеся, обычно свободноплавающие водоросли с цилиндрическими одноядерными клетками, с целлюлозной оболочкой, покрытые слизистым чехлом. Строение хроматофоров строго определенно и служит диагностическим признаком для каждого рода. Размножение осуществляется фрагментацией нити на отдельные короткие участки, апланоспорами и акинетами, конъюгация лестничная или боковая. Зигота прорастает одним проростком.
Большинство видов живут в стоячих или слабопроточных пресных водоемах. Порядок включает более 15 родов, около 700 видов, объединенных в 3 семейства: спирогировые, зигне- мовые и мужоциевые.
Род ctjupoiupa (Spirogyra) широко распространен в стоячих и медленно текущих водах, нередко образуя большие массы тины ярко-зеленого цвета.
Нити спирогиры, свободноплавающие или прикрепленные ризоидами к субстрату, состоят из длинных цилиндрических, расположенных в один ряд клеток, с хорошо заметной и довольно толстой оболочкой. Снаружи нити одеты слизистым чехлом.
Характерным признаком представителей данного рода является то, что пристенные хроматофоры их клеток имеют вид спирально изогнутой ленты, с городчатыми или рассеченными краями (рис. 1.62). По средней линии хроматофора или по его гребню расположены хорошо заметные пиреноиды.
Полость клетки заполнена большой вакуолью. В. центре клетки находится крупное ядра с^ядрышкода, заключенное в цитоплазматический мешочек и подвешенное на цитоплазма- тических тяжах.
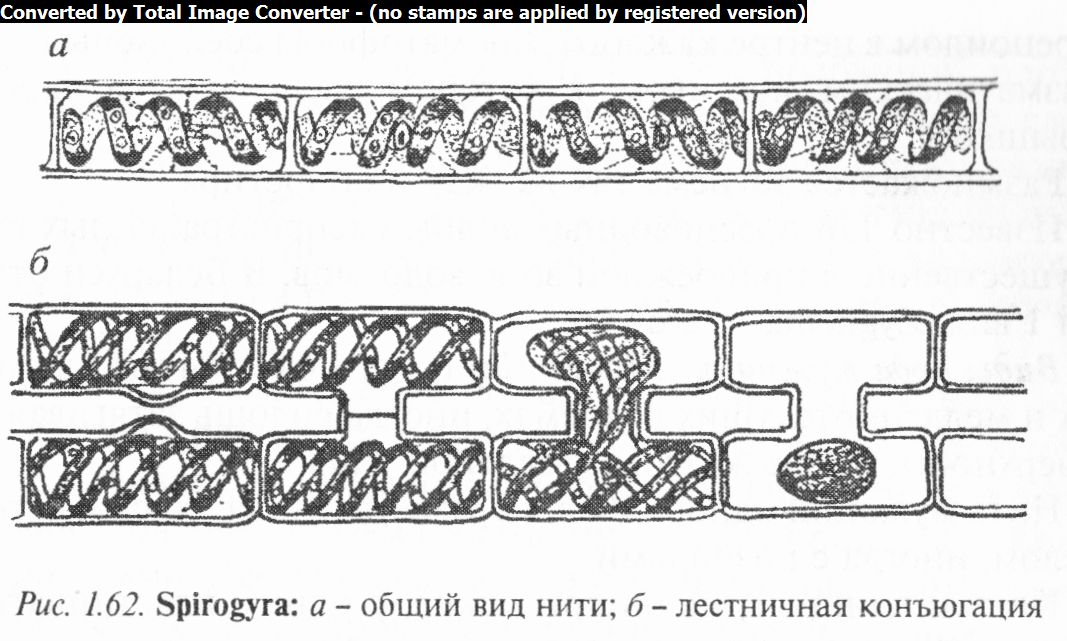
При половом размножении обычно две нити располагаются параллельно друг к другу и срастаются при помощи копуля- ционных отростков или мостиков. Оболочки в месте их соприкосновения растворяются и образуется сквозной канал, через который сжавшееся содержимое одной клетки перемещается в другую и сливается с ее протопластом. Кроме так называемой лестничной конъюгации изредка наблюдается и боковая: через боковой канал над поперечной перегородкой содержимое клетки переходит в соседнюю клетку той же нити. Зиготы, одетые толстой трехслойной оболочкой, после периода покоя прорастают. Этому предшествует редукционное деление ядра: из четырех получающихся гаплоидных ядер три отмирают, а одно остается ядром единственного проростка, выходящего через разрыв наружных слоев оболочки зиготы.
Известно около 340 видов, в том числе в Беларуси 8.
Широко распространены С. сжатая (S. condensata), С. речная (S. fluviatilis), С. толстая (S. crassa), С. простая (S. neglecta) и др.
Водоросли рода зигнема (Zygnema) (рис. 1.63) встречаются часто в тех же условиях, что и спирогира. Нити ее тоньше нитей спирогиры, с короткими цилиндрическими клетками, чаще всего одиночные, неветвящиеся. Отличительным признаком является наличие в клетках зигнемы симметрично расположенных двух крупных звездчатых осевых хроматофоров с
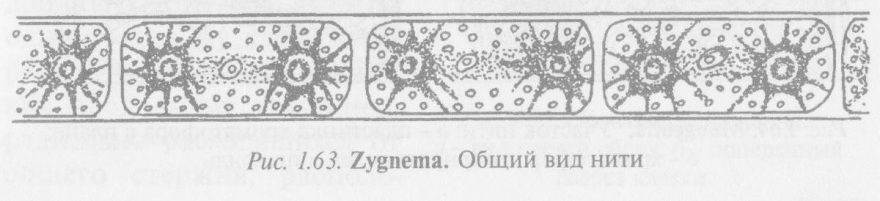
пиреноидом в центре каждого^Хроматофоры соединены цито- плазматическим мостиком, в котором располагается ядро с ядрышком.
Размножается зигнема так же, как и спирогира.
Известно 126 пресноводных видов, распространенных преимущественно в прибрежной зоне водоемов. В Беларуси отмечен 1 вид: Zygnema Agardh sp.
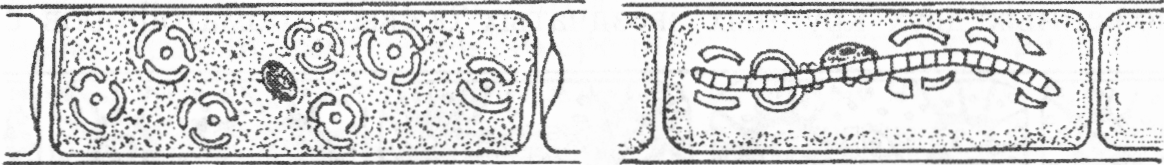
Виды рода мужоция (Mougeotia) (рис. 1.64) обитают в стоячих и медленнотекущих водоемах, иногда сплошь затягивая их поверхность в виде желтовато-зеленой тины.
Нить мужоции однорядная, неразветвленная, со слизистым чехлом, иногда с ризоидами.
Клетки длинные, цилиндрические, с одним пластинчатым хроматофором, расположенным в срединной плоскости клетки по длинной ее оси, с несколькими пиреноидами. Хроматофоры способны поворачиваться внутри клеток на 90°, что можно наблюдать при разной интенсивности освещения водорослей. При сильном освещении хроматофоры принимают вид узкой зеленой полоски, а при его снижении возвращаются в обычное положение и видны в клетке со стороны пластинки.
Иногда в клетке одна половинка хроматофора повернута ребром, а другая своей широкой стороной обращена к наблюдателю. Ядро расположено в центре и видно в тех клетках, где хроматофор повернут ребром.
Известно около 120 видов, из них 2 отмечены в Беларуси. Широко распространены М. лестничная (М scalaris), М. слизистая (М. gelatinosa), М. зеленая (М viridis).
Порядок Десмидиальные (Desmidiales). Он объединяет кок- коидные, преимущественно одноклеточные водоросли, реже колониальные в виде нитей или цепочек с резко выраженной двухсторонней симметрией клеток. У подавляющего большинства клетка разделена перетяжкой на две симметричные половинки; у немногих родов, не имеющих перетяжки, двухсторонняя симметрия отмечена в расположении и строении хромато-
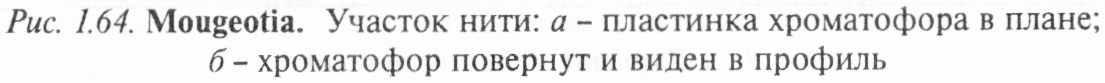
форов. Оболочка клеток состоит из двух половинок, плотно сомкнутых в плоскости симметрии, трехслойная и имеет слож- ноустроенный поровый аппарат. Хроматофоры имеют разную форму и строение, но всегда сохраняют симметрию полуклеток. Количество пиреноидов зависит от размеров хроматофоров и колеблется в пределах от 1 до 100. Ядро одно, располагается в перешейке.
Бесполое размножение осуществляется посредством деления клетки в плоскости симметрии, при этом каждая полуклетка достраивает недостающую половинку. Половой процесс — конъюгация между двумя клетками. Зигота прорастает двумя проростками. Десмидиевые водоросли способны формировать партеноспоры из непрокопулированных клеток, а также апла- но- и реже хламидоспоры.
Представители порядка — преимущественно пресноводные обитатели. Они предпочитают воду с кислой реакцией, бедную кальцием. Поэтому наиболее разнообразно и обильно они представлены в низинных сфагновых болотах. Многие из них живут в неглубоких лужах и торфяниках, на почвах, в горных озерах с чистой водой и др.
В порядке выделено 3 семейства: пениевые, клостериевые и космариевые с типичными одноименными родами и видами (около 4000).
Род кяостериум (Closterium) (рис. 1.65) объединяет виды, живущие одиночными клетками или иногда склеенные в пучки. Клетки у большинства видов изогнутые в форме полумесяца, реже прямые. Перетяжки посередине клетки нет. Оболочка состоит из двух половин, спаянных в плоскости симметрии, гладкая или с продольными штрихами. В_каж- дой полуклетке находится по одному осевому хроматофо- ру, состоящему из нескольких продольных пластинок,
радиально расходящихсяОТ а- вид клетки сооку; о - поперечныйобщего стержня, располо-
женного вдоль длинной оси хроматофора, пиреноиды либо в ряд, либо беспорядочно разбросаны от 1 до 20. Середина клетки, не занятая хроматофорами, бесцветна и представляет собой цитоплазматический мостик, в котором помещается крупное ядро. На концах клетки по 1 — 3 вакуоли и поры, выделяющие слизь, за счет которой осуществляется скользящее передвижение водоросли.
Известно более 300 видов (по некоторым сведениям 85), обитающих в пресноводных водоемах, 34 в Беларуси. Широко распространены в водохранилищах, реках, прудах, озерах и торфяных болотах К. ланцетовидный (С. lanceolatum), К. четко- носный (С. moniliferum), К. серповидный (С. lunula), К. малый (С. parvulum).
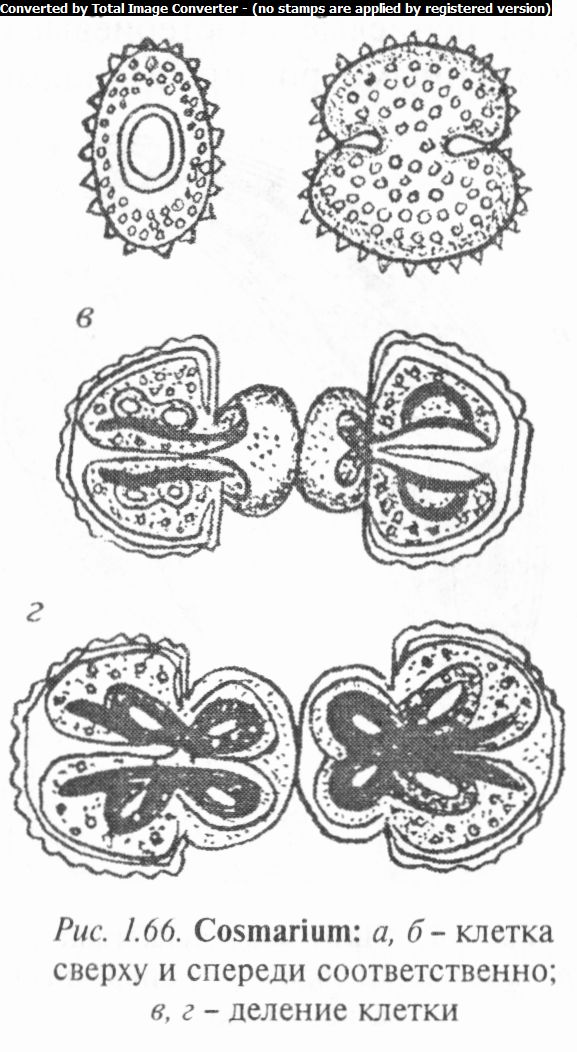
Род космариум (Cosmarium). Виды рода космариум имеют форму двух полукруглых или слегка удлиненных половинок, соединенных перетяжкой (рис. 1.66). В полуклетках располагаются по одному, реже по два осевых хроматофора. У некоторых видов хроматофоры постенные, лентовидные и имеют по одному пиреноиду. Ядро находится в перетяжке.
Обитают космариумы в планктоне рек, прудов, в болотах, предпочитают кислые водоемы с торфяным грунтом.
Известно около 800 видов и 62 в Беларуси. Повсеместно распространены К. пухловид- ный (С. subtumidum), К. чет- ковидный (С. inoniliforme), К. Менегини (С. meneghinii), К. зернистый (С. granatum) и др.
Род стаураструм (Stauras- truin) (рис. 1.67) включает большое количество видов одноклеточных водорослей с разнообразной формой клеток (3 — 5-угольной, у некоторых радиальной). Часто углы полуклеток образуют всевозможные выросты. Оболочка у большинства водорослей орнаментированная (грану-
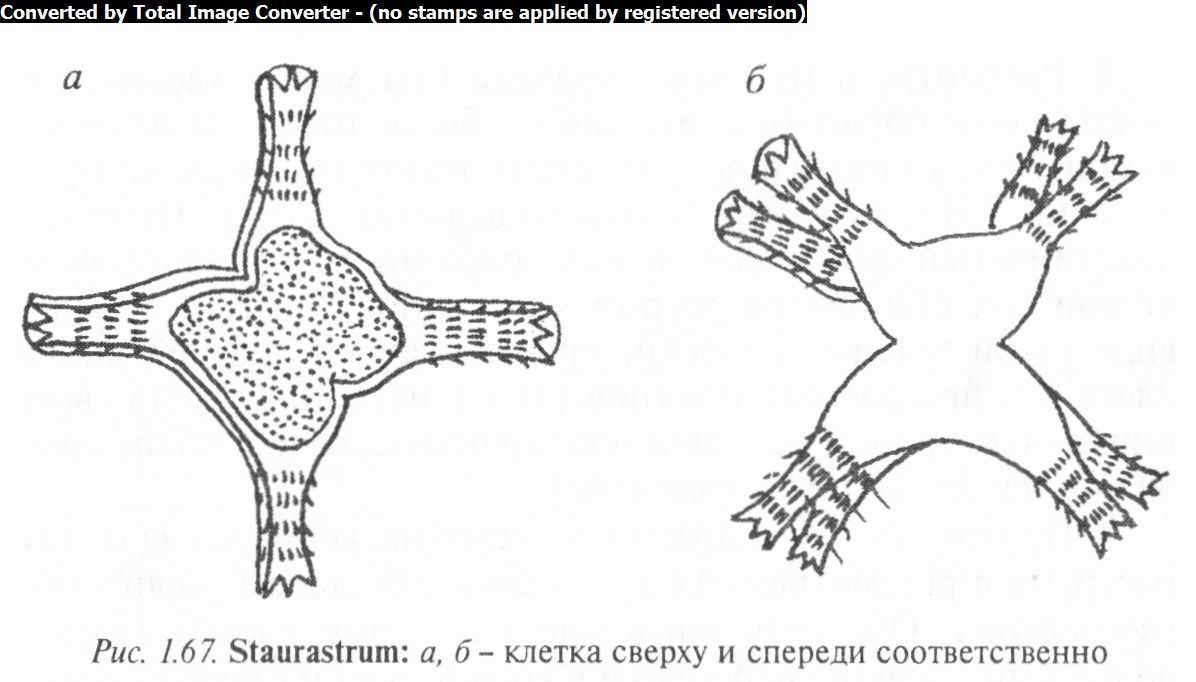
лы, шипы, зубчики и др.). Хроматофоры обычно осевые, по одному в полуклетке, с лопастями и одним пиреноидом, иногда постенные. Размножение преимущественно бесполое (поперечным делением в области перешейка) и половое — конъюгация.
Известно около 600 видов стаураструма и 21 в Беларуси. Они распространены в реках, озерах, прудах, среди планктона и обрастаний высших растений. Часто встречаются С. четырехрогий (S. tetracerum), С. тонкий (S. gracile), С. удивительный (S. paradoxum) и др.
Задания
1. Ознакомиться с внешним видом спирогиры. Обратить внимание на изумрудно-зеленый цвет и слизистую н<
Дата добавления: 2021-03-18; просмотров: 922;
Поиск по сайту
Узнать еще
- II. Види виробничої документації та порядок її ведення
- II. Порядок разработки, утверждения, внесения изменений в Инструкцию по делопроизводству
- III. Порядок присвоения спортивных званий
- IV. Движение поездов при неисправности электрожезловой системы и порядок регулировки количества жезлов в жезловых аппаратах
- IV. Порядок выполнения работы
- IV. Порядок присвоения спортивных разрядов
- V. Порядок лишения, восстановления спортивных званий
- VI. Порядок лишения, восстановления спортивных разрядов

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
