СИСТЕМАТИКА ВОДОРОСЛЕЙ
1.4. ОТДЕЛ ЦИАНОБАКТЕРИИ {CYAN О BACTERIA), ИЛИ СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ (CYANOPHYTA)
Цианобактерии, или синезеленые водоросли (или цианеи), представляют собой древнейшую уникальную в морфологическом и физиологическом отношении группу организмов. Многие свойства синезеленых водорослей (фиксация азота, прижизненные выделения органических веществ, оксигенный тип фотосинтеза) определяют их чрезвычайно важную роль в почве и водоемах. В последнее время цианеи стали объектами исследований биохимиков и физиологов, гидро- и микробиологов, генетиков и растениеводов, а также специалистов по космической биологии.
Отдел включает одноклеточные, колониальные и многоклеточные (нитчатые), от микроскопических до видимых простым глазом организмы различной морфологической структуры. Колониальные формы существуют на протяжении всей жизни или на отдельных стадиях развития водоросли. Многоклеточные цианеи живут отдельными нитями или собраны в дернинки. Они имеют симметричные или асимметричные, простые или разветвленные трихомы (тела), интеркалярную или апикальную зоны роста. У ряда нитчатых цианей имеются специализированные клетки — гетероцисты с сильно утолщенными бесцветными двухслойными оболочками. Они принимают участие в процессе фиксации атмосферного азота.
Клетка одета сложной по строению и составу пектиновой оболочкой, часто ослизняющейся, под которой располагается протопласт, как правило, лишенный вакуолей с клеточным соком. В клетке отсутствуют обособленное ядро, хроматофоры, аппарат Гольджи, митохондрии, эндоплазматическая сеть.
Цитоплазма делится на центральную часть — центроплазму (нуклеоплазму) и окрашенную периферическую - хромато- плазму. Строение центроплазмы — аналога ядра у синезеленых водорослей — близко к идентичным структурам бактериальных клеток и существенно отличается от строения клеток с оформленными ядрами.
В хроматоплазме расположены фотосинтезирующие ламел- лярные структуры и пигменты: хлорофилл а, каротиноиды и билихромопротеиды (синие фикоцианин и аллофикоцианин и красный фикоэритрин), поглощающие свет в области 540 — 630 нм, которая слабо используется всеми другими фотосинте- зирующими организмами (такой способностью обладают и красные водоросли). Благодаря уникальному и лабильному составу пигментов, цианеи способны к поглощению света различных длин волн.
Запасные вещества представлены гликогеном, волютином, цианофициновыми зернами. У многих синезеленых водорослей в цитоплазме имеются газовые вакуоли.
Размножаются цианеи простым бинарным делением клеток, распадом колоний, фрагментацией нитей на отдельные участки таллома — гормогонии, способные прорастать в новые талломы, а также гонидиями, кокками, планококками. Гони- дии — мелкие клетки со слизистой оболочкой, отделяющиеся от таллома или располагающиеся внутри эндоспор.
Кокки — одноклеточные фрагменты таллома, не имеющие четко выраженной оболочки.
Планококки — мелкие голые клетки, способные к самостоятельному движению.
Многие нитчатые цианеи образуют из одной, а иногда из двух и более соседствующих вегетативных клеток споры (аки- неты), которые служат главным образом для перенесения неблагоприятных условий. Спорообразование свойственно ностоковым и хамесифоновым, у последних образуются экзо- и эндоспоры, служащие для размножения.
Половой процесс и подвижные жгутиковые формы и стадии развития у цианобактерий не выявлены.
Синезеленые водоросли распространены в пресных и соленых водах, на поверхности почвы, скалах, в горячих источниках, входят в состав лишайников. Вместе с бактериями цианеи обогащают почву органикой и азотом, способствуют эвтрофи- рованию водоемов, являются кормом для зоопланктона и рыб, могут быть использованы для получения ряда ценных веществ, продуцируемых ими в процессе жизнедеятельности (аминокислоты, витамин В12, пигменты и др.) В период массового размножения в водоемах, так называемого «цветения» воды, некоторые цианеи токсичны для водных животных. Отдельные виды могут использоваться в пищу.
Синезеленые водоросли делят на 3 класса: Хроококкофици- евые, Хамесифонофициевые и Гормогониофициевые. Классификация основана на особенностях строения таллома и размножения цианей.
Класс Хроококкофициевые (Chroococcophyceae)
Класс включает колониальные и одноклеточные организмы. Колонии образуются в основном не разошедшимися после деления клетками, реже путем их слипания. Клетки в колонии располагаются преимущественно беспорядочно. Они не дифференцированы на основания и вершину. Размножаются вегетативным путем. Гетероцисты, а также эндо- и экзоспо- ры отсутствуют. В классе 2 порядка и 35 родов.
Порядок Хроококкаль- ные (Chroococcales). Объединяет широко распространенные одноклеточные и колониальные формы, свободноплавающие или лежащие на субстрате. Отдельные представители ведут прикрепленный образ жизни.
Род микроцистис (Microcystis) - это микроскопические, как правило, бесформенные комочки слизи, в которую погружены беспорядочно расположенные мелкие шаровидные клетки. У многих видов клетки под микроскопом кажутся почти черными из-за обилия в них газовых вакуолей, благодаря которым коло-
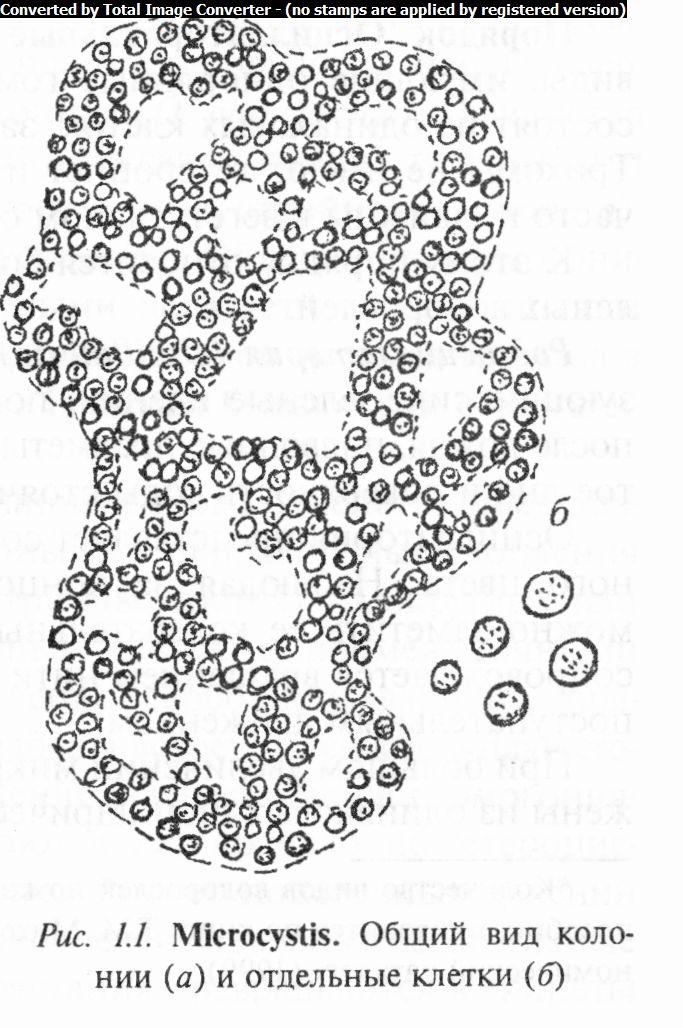
нии всплывают на поверхность воды. Очертания слизи этой колонии могут быть самыми разнообразными, причем иногда в слизи возникают своеобразные ячейки, благодаря чему колонии становятся сетчатыми (рис. 1.1).
Известно около 25 видов, распространенных в пресных и морских водоемах, а также в почве. В Беларуси выявлено 19 видов и 26 разновидностей*. Встречаются в водохранилищах, озерах и реках. Наиболее распространены М. синевато-зеленый {М. aeruginosa), М. Гревилля (М grevillei) и М. порошковидный (М pulverea). Некоторые виды токсичны.
Класс Гормогониофициевые (Hormogoniophyceae)
Класс объединяет многоклеточные водоросли нитчатой или трихомной формы, у которых протопласты соседних клеток взаимосвязаны плазмодесмами. Трихомы голые или покрытые слизистыми влагалищами. Многим из них свойственны гетероцисты. Размножение происходит гормогония- ми, реже акинетами. Класс насчитывает свыше 10 порядков. Наиболее важными из них являются осциллаториальные и ностокальные.
Порядок Осциллаториальные (Oscillatoriales). Включает виды, имеющие однорядные гомоцитные трихомы, которые состоят из одинаковых клеток, за исключением верхушечной. Трихомы не имеют гетероцист и почти всегда лишены спор, часто подвижны в вегетативном состоянии.
К этому порядку относится большинство нитчатых синезеленых водорослей.
Род осциллатория (Oscillatoria) включает виды, часто образующие сине-зеленые пленки, покрывающие влажную землю после дождя, подводные предметы и растения, затягивают илистое дно и поверхность воды стоячих водоемов.
Осциллатория представляет собой длинные нити сине-зеле- ного цвета. Наблюдая за концом нити под микроскопом, можно заметить ее колебательные движения. Это колебание сопровождается вращением нити вокруг собственной оси и ее поступательным движением.
При большом увеличении микроскопа видно, что нити сложены из одинаковых цилиндрических клеток, за исключениемверхушечных, которые по форме несколько отличаются от остальных (рис. 1.2).
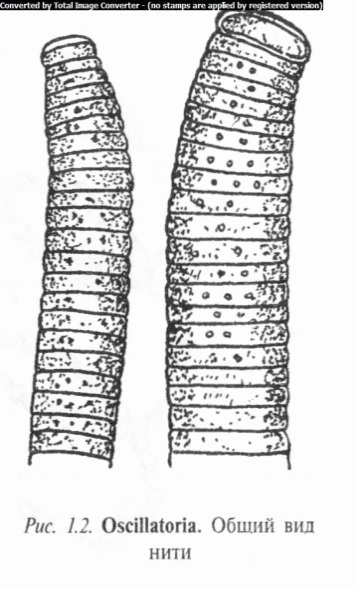
Внутри клетки можно видеть зернистые включения — цианофициновые зерна, располагающиеся, как правило, вдоль поперечных перегородок. Нить размножается путем распада на отдельные участки — гормогонии, вырастающие в новые нити.
Известно более 100 видов. В Беларуси выявлено 39 видов и 49 разновидностей. Обитают в бентосе и планктоне преимущественно пресных водоемов, иногда вызывая их «цветение». Прикрепляются к подводным предметам. Живут в иле, на
сыром песке или почве, а также встречаются в сточных водах. Наиболее распространены в планктоне прудов и озер: О. озерная (О. limnetica), О. планктонная (О. planctonica), О. илистая {О. limosa), О. тонкая (О. tenuis). На сваях, камнях, поверхности стоячих вод повсеместно встречается О. стройная (О. formosa).
Порядок Ностокальные (Nostocales). Объединяет гормогони- евые водоросли с гетероцитными неразветвленными нитями или нитями с ложным ветвлением (за счет прорыва трихома в сторону), часто с акинетами. Трихомы бывают как с влагалищами, так и без них.
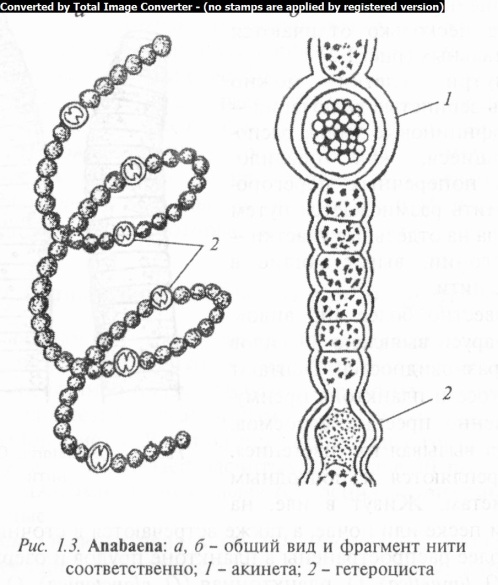
Род анабена (Anabaena) одноименного семейства представлен одиночными или собранными в неправильные скопления трихомами. Трихомы симметричны, состоят из округлых или бочонкообразных вегетативных клеток с промежуточными гетероцистами, преимущественно свободноплавающие, прямые или изогнутые. Виды анабены встречаются как в планктоне, так и в бентосе. Размножение осуществляется гормогония- ми, на которые нити распадаются, как правило, по гетероцис- там. Гормогонии растут только за счет поперечных делений клеток. Кроме того, у этих водорослей отдельные вегетативные клетки за счет сильного разрастания превращаются в акинеты (рис. 1.3). Акинеты значительно крупнее вегетативных клеток и выделяются своей яркой сине-зеленой окраской на почти черном от газовых вакуолей фоне остальных клеток. Содержимое акинет обычно зернистое, что в большинстве случаев обусловлено накоплением цианофициновых зерен. Известно около 100 видов, из них 28 в Беларуси. Встречаются в планктоне пресных вод, некоторые в солоноватых водах и на влажной почве. Наиболее распространены А. Гассаля (A. hassalii), А. Шереметьевой (A. scheremetievii), А. изменчивая (A. variabilis), А. спиралевидная (A. spiroides), А. «цветения» воды (A. flos-aquae) и др.
Род носток (Nostoc) характеризуется сложными слизистыми или студенистыми колониями разных размеров (от микроскопически мелких до крупных, достигающих размера сливы) и формы, часто сферической. В слизи находятся сложноперепле- тенные нити, похожие на нити анабены. Размножается посредством гормогониев. Они становятся подвижными и покидают материнскую колонию, слизь которой к этому времени расплывается. После некоторого периода движения гормогонии останавливаются, теряют газовые вакуоли (у бентосных видов) и прорастают в спирально извитые нити. Затем в результате многократных делений клеток гормогония продольными или косыми перегородками формируется зигзагообразная нить, свойственная ностокам. Эти нити покрываются обильной слизью и таким образом возникает молодая колония (рис. 1.4). Наблюдается и спорообразование, при котором многие вегетативные клетки превращаются в акинеты, обычно мало отличающиеся по форме и размерам от вегетативных клеток.
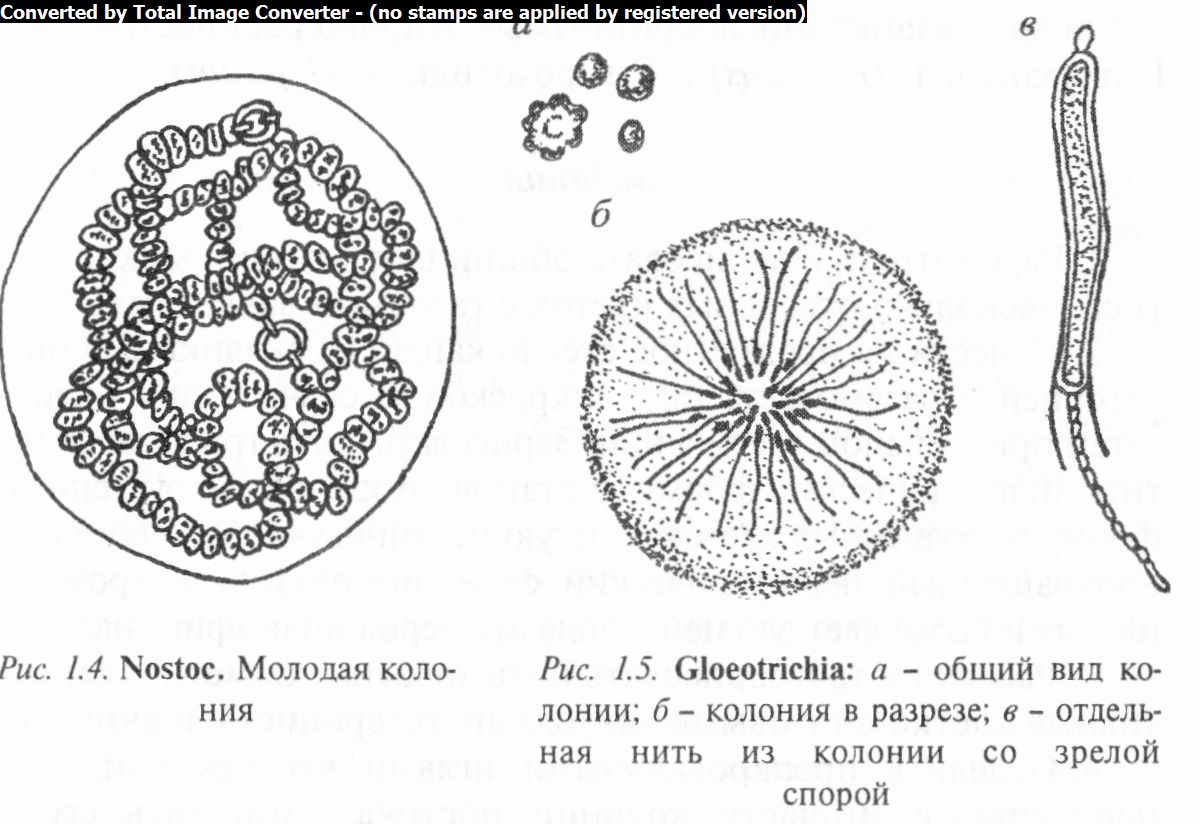
Виды ностока (более 50, в том числе 8 в Беларуси) широко распространены в водоемах и на почве. Некоторые виды являются съедобными. Типичный представитель рода — Н. сливо- видный (N. pruniforme), занесенный в Красную книгу Республики Беларусь.
Род глеотрихия (Gloeotrichia) включает виды, у которых нити соединяются общей слизью в шаровидные или полушаровидные колонии. Бичевидные асимметричные нити внутри слизи располагаются радиально, имеют расширенные концы, несущие гетероцисты и акинеты, обращенные внутрь колонии (рис. 1.5). Размножается посредством гормогониев. Известно 15 видов, в том числе 3 в Беларуси. Встречаются преимущественно в стоячих пресных водоемах; вначале прикреплены к субстрату, затем плавают свободно; из них только два вида — планктонные организмы. Широко распространены Г. плавающая (G. natans) и Г. гороховидная (G. pisum).
Задания
1. Рассмотреть и зарисовать общий вид колонии микроцис- тиса, несколько отдельных клеток с газовыми вакуолями.
2. Нанести на предметное стекло каплю из склянки с осцил- латорией и рассмотреть под микроскопом сначала при малом, затем при большом увеличении. Зарисовать часть трихома. Отметить цилиндрическую форму вегетативных клеток, закругленную форму верхушечных клеток, тонкую пектиновую оболочку, силь- ноокрашенный периферический слой цитоплазмы — хромато- плазму и более светлую центроплазму, зерна цианофицина.
3. Рассмотреть и зарисовать нить анабены. Отметить вегетативные клетки с газовыми вакуолями, гетероцисты и акинеты.
4. Отделить препаровальными иглами кусочек слизи из периферической части колонии ностока, поместить его в каплю воды на предметное стекло и рассмотреть под микроскопом. Зарисовать общий вид части колонии при малом увеличении и отдельную нить — при большом. Отметить вегетативные клетки и гетероцисты.
5. Рассмотреть и зарисовать колонию глеотрихии. Затем разрушить колонию, взять кусочек слизи, содержащей нити глеотрихии, и рассмотреть под микроскопом. Отметить, что гетероциста лежит в основании нити. Клетки, составляющие нить, по мере удаления от гетероцисты становятся тоньше и на вершине переходят в бесцветный волосок.
Вопросы для самоконтроля
1. Чем отличаются цианобактерии от фототрофных зеленых и пурпурных бактерий по строению тела, набору пигментов и типу фотосинтеза?
2. Чем отличается строение клетки синезеленых водорослей от строения клетки других организмов?
3. Какие формы организации таллома и размножения известны у цианеи?
4. Какие пигменты и запасные продукты отмечены в клетках синезеленых водорослей?
5. В чем заключается уникальность фотосинтезирующего аппарата синезеленых водорослей?
6. Каковы особенности строения и функции гетероцист и акинет?
7. Каково значение синезеленых водорослей в природе и народном хозяйстве?
1.5. ОТДЕЛ ЭВГЛЕНОВЫЕ ВОДОРОСЛИ (EUGLENOPHYTA)
Отдел включает микроскопические одноклеточные организмы, снабженные одним или двумя жгутиками и активно двигающиеся. Форма тела эвгленовых водорослей удлиненная, овальная, эллипсовидная или веретеновидная. Целлюлозной оболочки нет; ее роль выполняет наружный уплотненный слой цитоплазмы — пелликула. Те виды, у которых пелликула мягкая, эластичная, обладают способностью менять форму тела. У немногих водорослей есть наружный твердый панцирь, обычно пропитанный солями железа, не прилегающий плотно к протопласту. Число и форма хроматофоров различны. Они бывают веретеновидными, лентовидными, дисковидными, звездчатыми, пластинчатыми. Зеленый цвет эвгленовых водорослей обусловлен наличием хлорофиллов а и Ь. Кроме них присутствуют каротины и ксантофиллы. Запасной продукт — парамилон, производное глюкозы; он откладывается на наружных, выступающих из хроматофоров частях пиреноидов в виде скорлупок или в цитоплазме в виде мелких зерен.
На переднем конце эвгленовых водорослей находится углубление, часто называемое глоткой. Оно является выводным концом для системы сократительных вакуолей, в которых скапливается жидкость с растворенными продуктами обмена веществ.
Движение эвгленовых водорослей совершается за счет метаболических изменений формы тела и с помощью жгутика.
Размножение происходит продольным делением клетки пополам в подвижном или неподвижном состоянии. При неблагоприятных условиях у некоторых эвгленовых водорослей формируются покоящиеся цисты с толстыми оболочками. Половой процесс не доказан.
Эвгленовые водоросли — обычные обитатели небольших пресных стоячих водоемов, вызывающие при массовом развитии «цветение» воды. Этой группе растений свойственны все три основных типа питания: фототрофное, сапротрофное и голозойное (заглатывание оформленных частиц органического вещества или мелких организмов), иногда смешанное (мик- сотрофное).
Отдел включает 1 класс Эвгленофициевые.
Класс Эвгленофициевые (Euglenophyceae)
Класс объединяет несколько порядков, различия между которыми основаны главным образом на деталях строения жгутикового аппарата.
Представителями порядка Эвгленальные (Euglenales) могут служить роды эвглена, трахеломонас и факус.
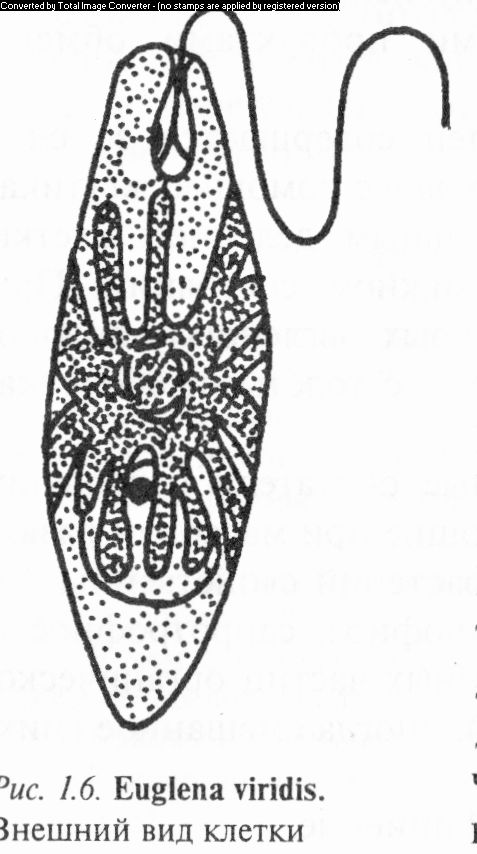
У водорослей рода эвглена (Euglena) клетки подвижные, веретено-, яйце- и лентовидные, цилиндрические, более или менее спирально закрученные. Передний конец суженный и закругленный, задний — заостренный, реже округлый или с узким шиловидным отростком (рис. 1.6). На переднем конце клетки имеются стигма, пульсирующие вакуоли и отверстие глотки, из которой выходит один из жгутиков, а второй, короткий, находится внутри глотки. Ядро одно, хроматофоров от одного до нескольких, с пиреноидами или без них.
Известно 155 видов, распространенных преимущественно в небольших пресных водоемах (лужи, озера, реки), болотах, на мокрой почве. Некоторые виды вызывают «цветение» воды зеленого или красного цвета. В Беларуси известно 25 видов. Часто встречаются Э. зеленая (Е. viridis), Э. спирогирная (Е. spirogyra), Э. игольчатая (£. acus), Э. хвостатая (Е. caudata) и др. Эвгленовые могут служить индикаторами качества воды.
включает в себя свободноплавающие организмы со жгутиком и твердым домиком. Строение домика является характерным признаком вида. Домики имеют разную форму, как правило, бурую окраску и спереди отверстие для выхода жгута (рис. 1.7). Стенки гладкие или с порами, сосочками, гранулами, шипами. Хроматофоры (два и более) зеленые, с пиреноидами или без них. Есть виды без хлорофилла — сапро- трофы. При размножении клетка делится внутри домика: одна из дочерних особей выскальзывает через отверстие наружу и вырабатывает новый домик.
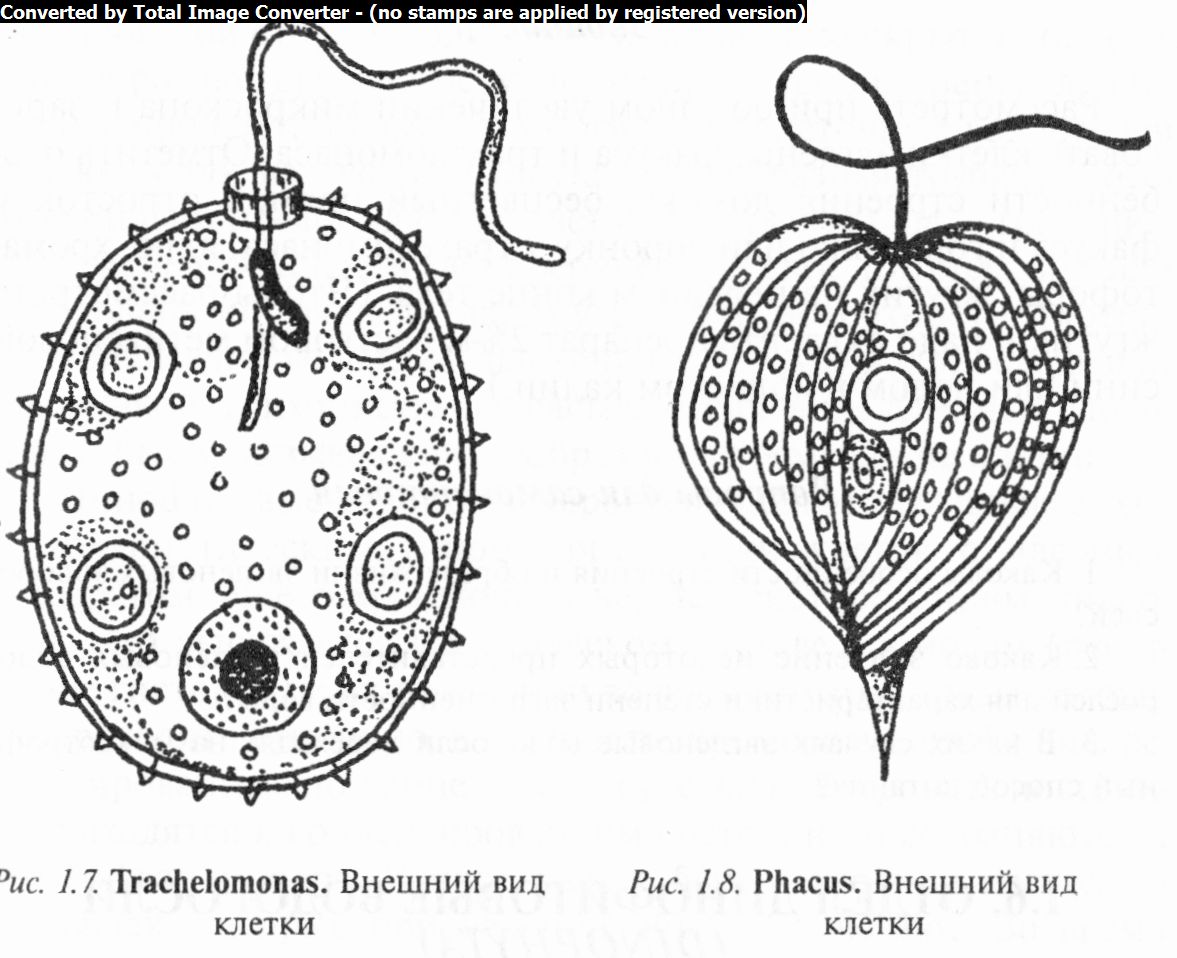
Известно около 200 видов, в Беларуси их 57 и 84 разновидности, распространенные в мелких водоемах с пресной водой. Наиболее известны Т. вольвоксовый (Т. volvocina), Т. мелкоще- тинковый (Т. hispida), Т. вооруженный (Т. armata), Т. продолговатый (Т. oblonga), Т. яйцевидный (Т. ovata) и др.
У видов рода факус (Phacus) (рис. 1.8) клетки плоскосжатые, более или менее штопоровидно скрученные, асимметричные, яйцевидные, овальные или шаровидные, с одним жгутиком, на заднем конце тела часто с бесцветным рулевым отростком. Пелликула плотная, бесцветная, со штрихами или рядами гранул, сосочков или шипиков. Хроматофоры многочисленные, мелкие, дисковидные, пристенные, без пиреноидов. Ядро одно (чаще в задней части клетки). В протопласте имеются зерна парамилона.
Известно около 140 видов, распространенных в мелких непроточных водоемах или в прибрежной части озер и рек, загрязненных органическими веществами. В Беларуси встречается 18 видов и 27 разновидностей. Наиболее распространены Ф. длиннохвостый (Ph. longicauda), Ф. мелкопузырчатый (Ph. vesiculosum), Ф. круглый (Ph. orbicularis), Ф. хвостатый (Ph. caudatus) и др.
Задание
Рассмотреть при большом увеличении микроскопа и зарисовать клетки эвглены, факуса и трахеломонаса. Отметить особенности строения домика, бесцветный прямой отросток у факуса и горлышко или воронку у трахеломонаса, ядро, хроматофоры, жгутик на переднем конце тела. (Чтобы рассмотреть жгутики, надо окрасить препарат 2%-м раствором метиленовой сини или иодом в иодистом калии.)
Вопросы для самоконтроля
1. Каковы особенности строения и образа жизни эвгленовых водорослей?
2. Каково значение некоторых представителей эвгленовых водорослей для характеристики степени загрязненности воды?
3. В каких случаях эвгленовые водоросли переходят на миксотроф- ный способ питания?
1.6. ОТДЕЛ ДИНОФИТОВЫЕ ВОДОРОСЛИ (DINOPHYTA)
Большинство динофитовых водорослей имеют монадное строение и представлены одиночными клетками. Реже встречаются амебоидные, пальмеллоидные, коккоидные и нитчатые формы. Для них характерна дорзовентральная форма тела: в строении клеток отчетливо выражены спинная и брюшная стороны, хорошо заметна разница между передним и задним концами тела.
Важным признаком для всех динофитовых является также наличие у клеток двух бороздок. Одна из них - поперечная - охватывает клетку по кольцу или спирали, но не смыкается полностью, другая - продольная — расположена на брюшной стороне клетки.
Клеточный покров у наиболее примитивных форм представлен гладким тонким перипластом (шаровидные формы). Большинство же динофитовых покрыты текой, состоящей из цито- плазматической мембраны, под которой в один слой располагаются компоненты теки - уплощенные пузыри (везикулы, цистерны), окруженные мембраной.
Два разных по длине, строению и даже функциям жгутика (один из них плавательный, другой — рулевой) прикрепляются на брюшной стороне; один почти полностью скрыт в поперечной бороздке, другой выступает из продольной и направлен по движению клетки назад. У многих одноклеточных водорослей имеются так называемая глотка (своеобразная трубка) и особые сильно преломляющие свет слизистые тельца — трихоцис- ты, расположенные в периферическом слое цитоплазмы или продольными рядами на внутренней поверхности глотки. Соприкасаясь во время движения с другой водорослью или песчинкой, трихоцисты выбрасываются в виде длинных слизистых нитей, вызывая скачкообразное перемещение клетки.
Динофитовые водоросли характеризуются так называемым мезокариотическим типом организации клеток с ядерным аппаратом, еще сохраняющим черты некоторой примитивности. Это выражается в химическом составе хромосом (отсутствие гистонов) и их поведении во время митоза, фазы которого проходят нетипично; в частности, хромосомы, слабо дифференцированные по длине из-за отсутствия центромер, постоянно находятся в конденсированном состоянии и сохраняются в интерфазном ядре. В ядре может быть одно или несколько ядрышек, которые при делении обычно исчезают. Во время митоза ядерная оболочка не исчезает и веретено деления не образуется.
В протопласте содержатся хроматофоры различной окраски (оливковые, бурые или коричневые, желтые, золотистые, красные, голубые, синие). Окраска обусловлена наличием хло- рофиллов а и с, каротинов а, [3, у, ксантофиллов (фукоксантин, перидинин, диадиноксантин, диатоксантин и др.). Имеются и бесцветные формы. Продуктами ассимиляции являются крахмал или масло, реже хризоламинарин, гликоген и др.
Только динофитовым водорослям присущи так называемые пузулы — области, похожие на вакуоли, соединяющиеся тонкими канальцами с окружающей средой и системой вакуолей. Они расположены обычно вблизи места отхождения жгутиков и выполняют роль осморегулятора.
Монадные формы размножаются делением клетки в подвижном состоянии на две части, каждая из которых позже вырабатывает недостающую половину. У других форм кроме деления клеток наблюдается образование зооспор, реже — апланоспор. Половой процесс в виде копуляции изогамет известен пока у немногих видов.
Динофитовые водоросли обитают в чистых пресных или соленых водоемах, встречаются в снегу. Среди них есть авто- и гетеротрофы. Наличие в загрязненных водоемах многих видов динофитовых является показателем сапробности при биологическом анализе воды. Некоторые формы участвуют в самоочищении воды.
Отдел Dinophyta делится на 2 класса: Динофициевые (Dinophyceae) и Десмофициевые (Desmophyceae).
Класс Динофициевые (Dinophyceae)
Класс объединяет организмы, имеющие целлюлозную оболочку (панцирь) или голые, одноклеточные или реже колониальные, в большинстве случаев подвижные. Встречаются также пальмеллоидные, ризоподиальные, коккоидные и нитчатые формы. Монадные клетки дорзовентральные, с поперечной и продольной бороздками, в которых располагаются жгутики, отходящие в месте пересечения бороздок. У многих представителей имеется стигма. Большинство видов являются фототрофными организмами, однако известны формы с голо- зойным питанием, а также паразиты и мутуалисты, обитающие в полости тела различных копепод.
В зависимости от типа организации таллома класс делится на ряд порядков. Типичными представителями класса являются роды перидиниум и церациум, относящиеся к порядку Перидиниальные.
Порядок Перидиниальные (Peridiniales). Объединяет формы с монадной организацией. Клетки многих представителей одеты мощным панцирем и имеют ясно выраженное дорзовент- ральное строение. У других видов отсутствует клеточная стенка и панцирь (они одеты перипластом).
Род перидиниум (Peridinium) (рис. 1.9) включает виды, имеющие шаровидный или яйцевидный коричневый панцирь, который состоит из постоянного числа пластинок. На поверхности панциря отчетливо выражены поперечная и продольная бороздки. Первая опоясывает спинную выпуклую сторону клетки, сходясь или расходясь обоими концами на брюшной плоской или вогнутой стороне. Она делит клетку на две примерно равные части: переднюю (апикальную) и заднюю (анти- пикальную). Продольная бороздка перпендикулярна к поперечной и проходит по брюшной стороне половины клетки, лишь частично заходя и на переднюю половину. От места пересечения обеих ложбинок отходят жгутики. Щитки соединены между собой поперечно-полосатыми швами, за счет расширения которых происходит рост панциря.
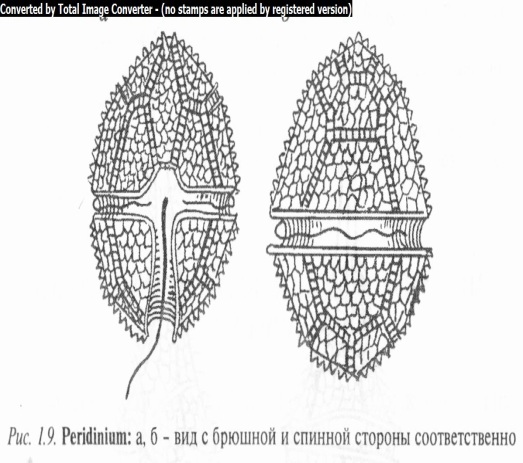
В протопласте имеются крупное ядро и дисковидные хроматофоры. Пиреноиды отсутствуют.
Размножение осуществляется делением клетки на две части или с помощью зооспор.
Известно около 200 видов, распространенных в пресных, солоноватых и морских водах (отдельные виды вызывают «свечение» моря). В Беларуси 9 видов. Чаще других встречаются П. опоясанный (P. cinctum), П. палатинский (P. palatinum), П. болотный (P. palustre), П. Вилле (P. willei) и др.
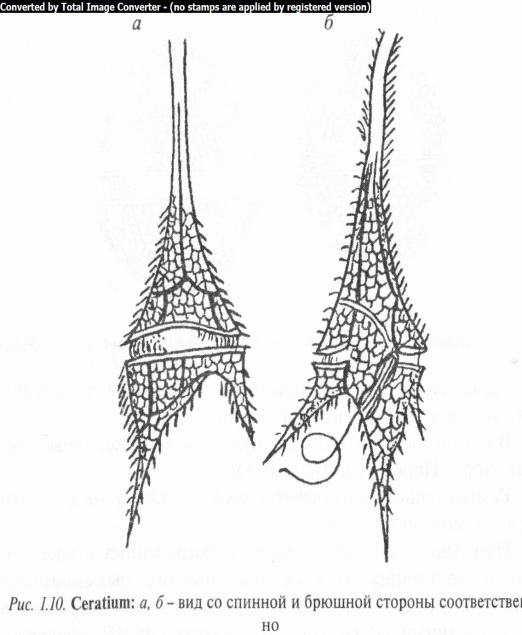
Виды рода церациум (Ceratium) (рис. 1.10) характеризуются наличием выростов или рогов: одного длинного переднего и 2 — 3 коротких задних. Поперечная бороздка окружает все тело церациума в самом широком его месте, продольная начинается от поперечной и идет вниз.
Клетки подвижные, двухжгутиковые, сильно вытянутые в продольном направлении и сплюснутые в спинно-брюшном. Спинная сторона выпуклая, брюшная вогнутая. Протопласт четко разделен на экто- и эндоплазму. Хроматофоры многочисленные, дисковидные.
Размножение осуществляется делением клетки вместе с панцирем. Обычно в конце вегетационного периода наблюдается цистообразование. У церациума рогатого половой процесс анизогамный.
Известно около 80 видов, среди которых 3 пресноводные, остальные — морские. В Беларуси описаны Ц. рогатый (C. cor nutum),
Ц. ласточковый (С. hirundinella). Могут вызывать «цветение» воды, придавая ей буровато-белесую окраску.
Задание
Рассмотреть и зарисовать клетки перидиниума и церациума со спинной и брюшной стороны при большом увеличении микроскопа. На рисунке отметить форму, размеры и окраску водорослей, особенности строения щитков, слагающих панцирь, наличие поперечной и продольной бороздок и двух жгутиков.
Вопросы для самоконтроля
1. Каковы особенности строения клетки динофитовых водорослей?
2. Для динофитовых водорослей характерно наличие уникального клеточного ядра — динокариона или мезокариона. Каковы особенности строения и функционирования ядра мезокариотического типа?
3. Какие фотосинтезирующие пигменты и продукты ассимиляции характерны для данной группы водорослей?
4. По каким признакам динофитовые водоросли сходны с растениями и по каким с животными?
5. Как у данной группы водорослей осуществляется размножение?
6. Каково значение динофитовых водорослей в природе и хозяйственной деятельности человека?
1.7. ОТДЕЛ КРИПТОФИТОВЫЕ ВОДОРОСЛИ (CRYPTOPHYTA)
Представители данного отдела - почти исключительно одноклеточные организмы монадной структуры, редко коккоид- ного или пальмеллоидного типа. Клетки бывают яйцевидными, эллипсовидными, веретеновидными, грушевидными и др. Они имеют обычно выпуклую спинную и вогнутую брюшную стороны.
Клетка одета перипластом и несет одну бороздку, идущую косо от небольшой выемки на переднем как бы срезанном конце клетки или поперек клетки, иногда по спирали. У некоторых представителей есть глотка. Два лентовидных жгутика с мастигонемами отходят от переднего конца клетки или сбоку.
Ядро эукариотического типа, одно, расположено в задней части клетки или, реже, посередине, имеет ядрышко.
У крипгофитовых водорослей обнаружена уникальная органелла — нуклеоморф. Она имеет сходство с ядром, находится в перипластидном пространстве, окружена двойной мембраной и содержит гранулярный матрикс. Предполагается, что наличие нуклеоморфа свидетельствует об эндосимбио- тическом происхождении хроматофоров криптофитовых водорослей.
Хроматофоры окрашены в разные цвета от сине-зеленого и оливково-зеленого до желто-бурого, бурого и красного. Окраска обусловлена наличием хлорофиллов а и с, каротиноидов (Р- и е-каротин, зеаксантин, диатоксантин, а также синий фикоцианин и красный фикоэритрин). Строма хроматофора содержит обычно вздутые тилакоиды, расположенные по два (иногда по три).
Продукт ассимиляции — крахмал, который откладывается в цитоплазме. У некоторых представителей запасными продуктами являются масло и хризоламинарин.
Питание фототрофное или сапротрофное, реже голо- зойное.
Размножение криптофитовых происходит путем продольного деления клетки надвое. При размножении оба жгутика отходят к одной клетке, а другая образует их заново. Половой процесс неизвестен.
Большинство криптофитовых живет в поверхностных, хорошо прогревающихся слоях воды. Они преобладают в небольших стоячих водоемах, лужах, различных прудах.
Отдел Криптофитовые водоросли включает 1 класс — Крип- тофициевые (Cryptophyceae) и один порядок — Криптомона- дальные (Cryptomonadales). Наиболее известным представителем порядка является род криптомонас.
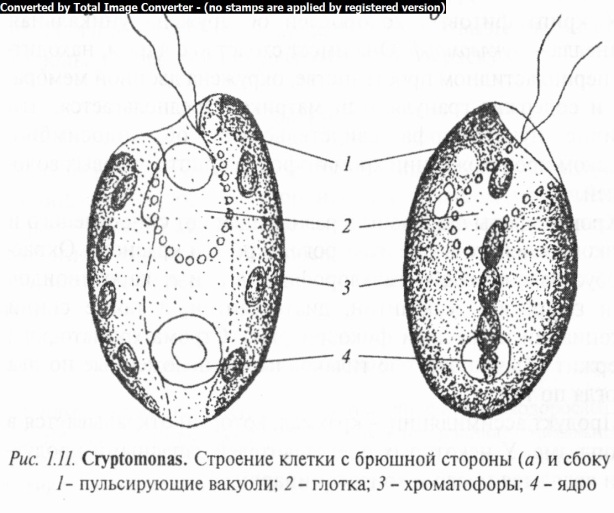
Род криптомонас (Cryptomonas) включает в себя подвижные одноклеточные водоросли с выпуклой спинной и плоской брюшной сторонами. Перипласт кожистый, часто продольно исчерченный. Передний конец клетки косо срезанный, с выемкой и косой бороздкой, которая переходит в глотку (рис. 1.11). От боковой стенки бороздки отходят два неравных жгутика. Ядро одно, пульсирующих вакуолей 1 — 3.
Известно более 50 видов, около 10 из них в Беларуси. Наиболее часто встречаются К. Марсона (С. marsonii), К. яйцевидный (С. ovata), К. изогнутый (С. reflex reflexa), К. обгрызенный (С. erosa) и др. Многие из них вызывают «цветение» воды.
Задание
Рассмотреть и зарисовать клетку криптомонаса с брюшной стороны и сбоку при большом увеличении микроскопа. Обратить внимание на форму, размеры и окраску водоросли, особенности строения перипласта, наличие на переднем конце косо срезанной клетки выемки с косой бороздкой, которая переходит в глотку. На рисунке отметить хроматофоры, глотку, жгутики.
Вопросы для самоконтроля
1. Каковы особенности строения клетки криптофитовых водорослей?
2. Какие пигменты обусловливают окраску таллома криптофитовых?
3. Какие продукты ассимиляции характерны для этой группы водорослей?
4. Как осуществляется размножение криптофитовых?
5. В чем выражается сходство и различие между динофитовыми и криптофитовыми водорослями по строению клетки, составу пигментов и продуктов ассимиляции и другим признакам?
1.8. ОТДЕЛ ЗОЛОТИСТЫЕ, ИЛИ ХРИЗОФИТОВЫЕ (CHRYSOPHYTA) ВОДОРОСЛИ
Золотистые водоросли характеризуются большим морфологическим разнообразием. Среди них встречаются амебоидные, пальмеллоидные, монадные, коккоидные, нитчатые, разнонит- чатые и даже пластинчатые формы. В большинстве своем они являются одноклеточными, но могут быть колониальными и многоклеточными. Клетки у одних водорослей голые или покрыты перипластом и способны к метаболическим изменениям формы тела, у других одеты оболочкой из целлюлозы и пектиновых веществ. У некоторых представителей клетки покрыты панцирем, состоящим из кремниевых чешуек (иногда с шипами), или заключены в домики. Движение осуществляется при помощи 1 — 2, иногда 3 — 4 жгутиков, одинаковой или разной длины.
Золотисто-желтая, желто-зеленая или бурая окраска зависит от хроматофоров (одного или нескольких), расположенных в постенном слое цитоплазмы и имеющих корытовидную форму. В хроматофорах содержатся хлорофилл а н с, кароти-ноиды ф-каротин) и ксантофиллы (желтый лютеин и буроватый фукоксантин, а также антераксантин, зеаксантин, неоксан- тин и виолаксантин). Запасные продукты — хризоламинарин (лейкозин) и липиды, откладывающиеся в цитоплазме. Ядро одно. У отдельных хризофит имеются 1 — 2 пульсирующие вакуоли и стигма, расположенная в хроматофоре. Ламеллы хроматофоров составляют пачки из трех или четырех тилакоидов. Тип питания большинства видов автотрофный и только у некоторых - гетеротрофный.
Бесполое размножение осуществляется делением клетки на две или распадом таллома на части, а также одно-, двухжгути- ковыми зооспорами, амебоидными клетками, апланоспорами. У некоторых видов описан половой процесс по типу изо-, холо- или автогамии. В результате полового процесса и в неблагоприятных условиях образуются цисты с толстой окремневшей оболочкой. После периода покоя циста прорастает, образуя зооспоры.
Большинство хризофитовых — планктонные водоросли чистых, не загрязненных органическими веществами пресных водоемов. Некоторые живут в морях или ведут эпифитный образ жизни.
Отдел Золотистые водоросли делят на 2 класса — Гетерохри- зофициевые (Heterochrysophyceae) и Изохризофициевые
(Isoch rysophyceae).
Класс Гетерохризофициевые (Heterochrysophyceae)
К данному классу относят представителей с различными типами организации таллома — монадные, ризоподиальные, пальмелловидные, коккоидные и нитчатые. В пределах класса выделяют 7 порядков: Хризомонадальные (Chrysomonadales), Кокколитальные (Coccolithales), Диктиохальные (Dictyochales), Ризохризидальные (Rhizochrysidales), Хризокапсальные (Chryso- capsales), Хризосферальные (Chrysosphaerales), Феотамниальные (Phaeothamniales). Деление на порядки основано на различиях в типе структуры таллома, числе и строении жгутиков, особенностях строения скелета и панциря, на наличии или отсутствии слизистой обвертки и т.п.
Порядок Хризомонадальные (Chrysomonadales). Самый многочисленный порядок, включающий водоросли монадной структуры, одноклеточные и колониальные, снабженные одним или двумя жгутиками часто неравной длины. Клеткиили голые, покрытые лишь плазмалем- мой, или же имеют целлюлозные футляры (домики) либо покрыты кремнистыми пластинками.
Большинство хризомонадальных водорослей — типичные активные планк- теры, и лишь некоторые ведут прикрепленный образ жизни.
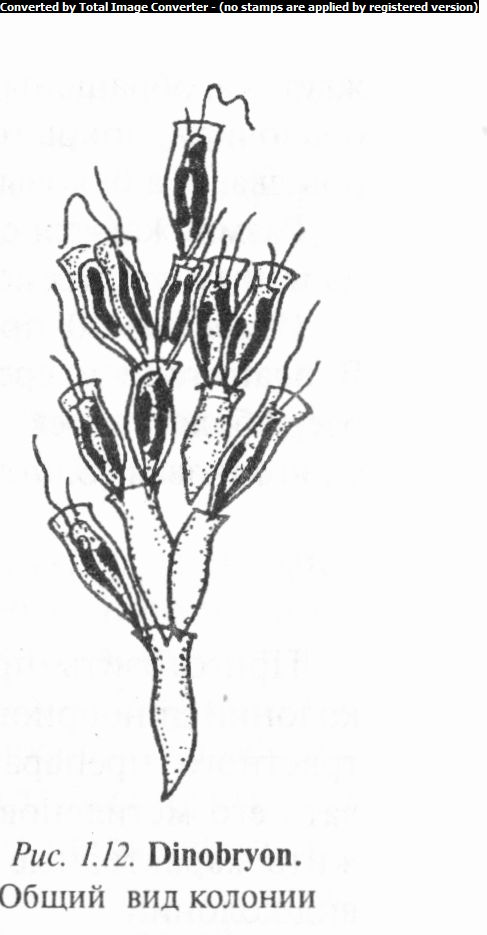
Род дииобрион (Dinobryon) включает одиночные или собранные в кустистые колонии организмы, свободно плавающие или прикрепленные к субстрату (рис. 1.12). Клетки динобриона прикреплены внутри бокаловидных прозрачных целлюлозных домиков при помощи сократительных стебельков. Из устья домика выставляются два жгутика неравной длины — длинный перистый и короткий боковой гладкий. Хроматофоров один-два, ядро одно. При размножении протопласт делится продольно, причем один из дочерних протопластов обычно остается в материнском домике, а другой выходит наружу и тут же формирует свой собственный. Клетки, сидящие у выхода из домика, могут превращаться в золотисто-оранжевые цисты, закрывающие отверстия домиков. В результате ряда таких делений возникают разветвленные колонии. Половой процесс — хологамия.
Известно более 20 видов, в том числе 12 в Беларуси. В реках, озерах, прудах и болотах часто встречаются Д. общественный (D. sociale), Д. цилиндрический (Д cylindricum), Д. спиральный (Д spirale), Д. расходящийся (Д divergens) и др.
Род синура (Synura) представлен свободноплавающими сферическими колониями, лишенными общей слизистой обертки (рис. 1.13). Клетки шаро- или обратнояйцевидные, расположены ради- ально, в центре колонии соединяются оттянутыми задними концами, а передние, несущие по два неравной длины
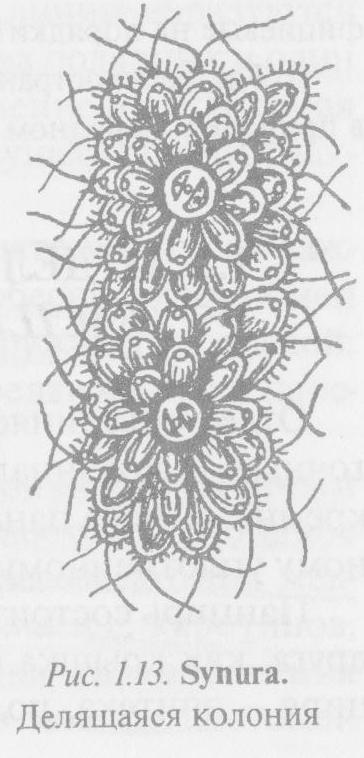
жгутика, обращены кнаружи. Каждая клетка одета
Дата добавления: 2021-03-18; просмотров: 1394;
Поиск по сайту
Узнать еще
- Б) Экологические особенности почвенных водорослей
- Значение водорослей
- Значение водорослей в природе и их использование человеком
- МЕТОДЫ СБОРА, ХРАНЕНИЯ И ИЗУЧЕНИЯ ВОДОРОСЛЕЙ
- Морфология и систематика микроорганизмов
- Морфология строение клеток и систематика
- ОБЩИЕ КРИТЕРИИ ДИАГНОСТИКИ. СИСТЕМАТИКА ПСИХОГЕННЫХ ЗАБОЛЕВАНИЙ
- Основные жизненные циклы водорослей

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
