Рецепторы химических посредников: типы, лиганд-рецепторные взаимодействия и механизмы регуляции
В предыдущих публикациях было описано, что несколько классов химических посредников обеспечивают передачу сигнала между клетками. К таким посредникам относятся нейромедиаторы и паракринные вещества, сигналы которых передаются быстро и на короткие расстояния. Другие посредники, например гормоны, распространяются на большие расстояния и, в некоторых случаях, медленнее. Независимо от типа химического посредника, клетка-мишень должна обладать механизмом обнаружения присутствия сигнала. После обнаружения сигнала запускается процесс преобразования этого сигнала в физиологически значимую реакцию, такую как деление клеток в ответ на ростовые стимулы.
Первым этапом действия любого межклеточного химического мессенджера является его связывание со специфическими белками клетки-мишени, называемыми рецепторами (или рецепторными белками). В общих биохимических терминах химический мессенджер функционирует как лиганд, а рецептор содержит сайт связывания, специфичный для данного лиганда. Связывание мессенджера с рецептором вызывает конформационные изменения (изменение третичной структуры; см. рис. 2.17) в рецепторе, активируя его. Эта активация инициирует последовательность клеточных событий, приводящих к ответу клетки, — процесс, называемый сигнальной трансдукцией. Термин «сигнал» относится к активации рецептора, а «трансдукция» описывает преобразование стимула в клеточный ответ. В данном разделе рассматриваются общие характеристики рецепторов, описываются взаимодействия рецептор-лиганд и приводятся примеры регуляции рецепторов.
Типы рецепторов. Рецепторы, связывающие межклеточные химические посредники, представляют собой белки или гликопротеины, расположенные либо на плазматической мембране клетки, либо внутри клетки (в цитозоле или ядре). Плазматическая мембрана является наиболее распространённым местом локализации, поскольку многие мессенджеры растворимы в воде и не могут проникать через богатый липидами (гидрофобный) бислой мембраны. Напротив, небольшое количество жирорастворимых мессенджеров способно проходить через мембраны и связываться со своими внутриклеточными рецепторами.
Рецепторы плазматической мембраны (показаны на рис. 5.1а) представляют собой трансмембранные белки, пересекающие всю толщину мембраны. Подобно другим трансмембранным белкам, такой рецептор имеет гидрофобные сегменты внутри мембраны, один или несколько гидрофильных сегментов, выходящих во внеклеточную жидкость, и дополнительные гидрофильные сегменты, выступающие во внутриклеточную среду. Поступающие химические мессенджеры связываются с внеклеточными участками рецептора, тогда как внутриклеточные участки участвуют в процессах передачи сигнала.
Внутриклеточные рецепторы (показаны на рис. 5.1б) расположены не в мембранах, а находятся либо в цитозоле, либо в ядре клетки и имеют совершенно иную структуру. Однако, как и рецепторы плазматической мембраны, они содержат сегмент, связывающий мессенджер, а также другие сегменты, функционирующие как регуляторные участки. Кроме того, внутриклеточные рецепторы обладают ДНК-связывающим сегментом, который отличает их от мембранных рецепторов. Это ключевое различие: рецепторы плазматической мембраны могут передавать сигналы без взаимодействия с ДНК, тогда как все внутриклеточные рецепторы передают сигналы посредством взаимодействия с генами.
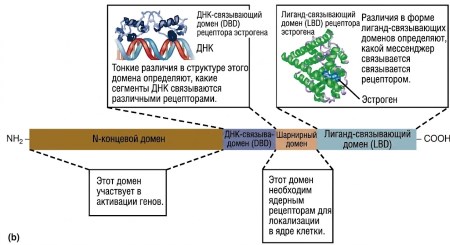
Рисунок 5.1. Два основных класса рецепторов химических посредников. (а) Структура типичного трансмембранного рецептора. Семь кластеров аминокислот, встроенных в фосфолипидный бислой, представляют собой гидрофобные участки альфа-спирали белка (показаны здесь в виде цилиндров). Обратите внимание, что место связывания гормона включает несколько сегментов, которые проникают во внеклеточную жидкость. Части внеклеточных сегментов могут быть связаны с углеводами (CHO). Аминокислоты, обозначенные черными кружками, представляют некоторые участки, в которых внутриклеточные ферменты могут фосфорилировать рецептор и тем самым регулировать его работу. (б) Схематическое изображение структурных особенностей типичного ядерного рецептора. Фактические структуры сегментов этих рецепторов известны и показаны здесь для рецептора человеческого эстрогена (стероидного гормона). (Примечание: Сегменты ядерных рецепторов, выполняющие различные функции, известны как «домены».)
Взаимодействия между рецепторами и лигандами. Взаимодействие между рецепторами и их лигандами определяется четырьмя основными характеристиками: специфичностью, аффинностью, насыщением и конкуренцией (терминологический словарь рецепторов приведён в таблице 5.1).

Специфичность относится к связыванию химического вещества со своим рецептором, которое инициирует события, приводящие к клеточному ответу. Существование рецепторов объясняет важнейшую характеристику межклеточной коммуникации — специфичность. Хотя данный химический мессенджер может контактировать со многими различными типами клеток, он воздействует только на определённые типы, поскольку клетки различаются по набору рецепторов. Только определённые типы клеток — иногда только один — экспрессируют конкретный рецептор, необходимый для связывания данного химического мессенджера (рис. 5.2).
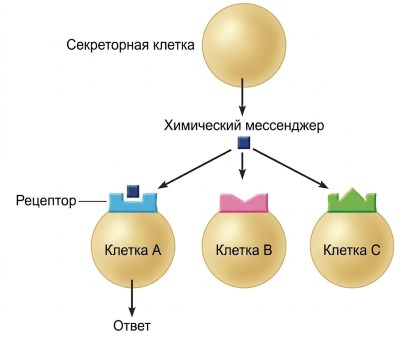
Рисунок 5.2. Специфичность рецепторов к химическим мессенджерам. Только клетка А имеет соответствующий рецептор для этого химического мессенджера; следовательно, она единственная в группе, которая является клеткой-мишенью для данного мессенджера.
Даже если разные типы клеток обладают рецепторами к одному и тому же сигналу, их реакция на этот сигнал может существенно различаться. Например, нейромедиатор норадреналин вызывает сокращение гладкой мускулатуры определённых кровеносных сосудов, но с помощью рецепторов того же типа подавляет секрецию инсулина поджелудочной железой. Таким образом, рецептор функционирует как молекулярный переключатель, который вызывает реакцию клетки при активации посредством связывания мессенджера. Аналогичным образом, идентичные переключатели могут управлять светом или радио, и один и тот же тип рецептора может вызывать разные реакции в разных типах клеток.
Аффинность определяет степень, в которой конкретный мессенджер связывается со своим рецептором (см. рисунок 5.3). Рецептор с высокой аффинностью будет связывать лиганд при более низких концентрациях мессенджера, чем рецептор с низкой аффинностью (см. рисунок 3.36). Различия в сродстве рецепторов к своим лигандам имеют важное значение для разработки терапевтических препаратов: рецепторам с высоким сродством требуется гораздо меньшее количество лиганда (меньшая доза) для активации.
Насыщение рецепторов (описанное в главе 3 на примере связывания лиганда с белком) в полной мере применимо и в данном случае (см. рис. 5.3). Реакция клетки на мессенджер возрастает по мере увеличения концентрации внеклеточного мессенджера, поскольку увеличивается число занятых рецепторов. Однако существует верхний предел этой чувствительности, так как доступно ограниченное количество рецепторов, и в какой-то момент они становятся полностью насыщенными.
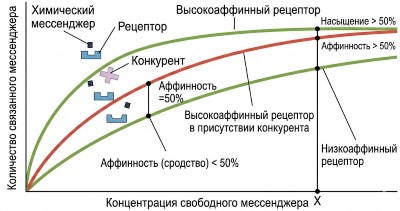
Рисунок 5.3. Характеристики рецепторов, связывающихся с посредниками. Рецепторы с высокой аффинностью будут иметь больше связанных посредников при заданной концентрации посредника (например, концентрации X). Присутствие конкурента снижает количество связываемого мессенджера до тех пор, пока при очень высоких концентрациях рецепторы не будут насыщены мессенджером. Обратите внимание на иллюстрацию, что в данном случае низкоаффинный рецептор имеет несколько иную форму в области, связывающей лиганд, по сравнению с высокоаффинным рецептором. Также обратите внимание на сходство некоторых частей форм природного мессенджера и его конкурента.
Конкуренция относится к способности молекулы конкурировать с природным лигандом за связывание с его рецептором. Конкуренция обычно возникает между посредниками, имеющими структурное сходство, и лежит в основе действия многих лекарственных средств (см. рис. 5.3). Когда исследователи или врачи хотят повлиять на действие конкретного мессенджера, они могут ввести конкурирующие молекулы, достаточно похожие по структуре, чтобы связываться с рецепторами этого мессенджера. Однако эти конкурирующие молекулы достаточно сильно отличаются от нативного лиганда, чтобы не активировать рецептор, тем самым блокируя связывание эндогенного мессенджера без индукции передачи сигнала.
Общий термин для соединения, блокирующего действие химического посредника, — антагонист; когда антагонист действует, конкурируя за место связывания, он известен как конкурентный антагонист. Одним из примеров служат блокаторы бета-адренорецепторов (также называемые бета-блокаторами), используемые при лечении высокого кровяного давления. Бета-блокаторы конкурируют с адреналином и норадреналином за связывание с бета-адренорецепторами. Поскольку адреналин и норадреналин обычно повышают кровяное давление (глава 12), бета-адреноблокаторы снижают кровяное давление, действуя как конкурентные антагонисты. Антигистаминные препараты — другой пример, полезный при лечении аллергических симптомов, вызванных избыточной секрецией гистамина тучными клетками (глава 18). Антигистаминные препараты являются конкурентными антагонистами, которые блокируют связывание гистамина с его рецепторами, предотвращая аллергические реакции.
Некоторые препараты, конкурирующие с природными лигандами за определённый тип рецепторов, действительно активируют рецептор и запускают клеточный ответ точно так же, как если бы связался эндогенный химический мессенджер. Такие препараты, известные как агонисты, используются в терапевтических целях для имитации действия мессенджера. Например, обычные противоотечные средства фенилэфрин и оксиметазолин (содержащиеся во многих назальных спреях) имитируют действие адреналина на альфа-адренорецепторы в кровеносных сосудах. При активации альфа-адренорецепторов гладкая мускулатура воспалённых расширенных кровеносных сосудов носа сокращается, что приводит к сужению сосудов, помогает открыть носовые проходы и уменьшить утечку жидкости.
Регуляция работы рецепторов. Рецепторы сами по себе являются объектом физиологической регуляции. Количество рецепторов, которыми обладает клетка, или аффинность рецепторов к их специфическому мессенджеру может быть увеличено или уменьшено в определённых системах. Важным примером является понижающая регуляция: когда в течение длительного времени поддерживается высокая концентрация внеклеточного мессенджера, общее количество рецепторов клеток-мишеней для этого мессенджера может уменьшаться. Понижающая регуляция снижает чувствительность клеток-мишеней к частой или интенсивной стимуляции, представляя собой локальный механизм отрицательной обратной связи.
Понижающая регуляция происходит из-за непрерывного синтеза и деградации рецепторов. Основным механизмом снижения количества рецепторов плазматической мембраны является интернализация. Связывание мессенджера с его рецептором может стимулировать интернализацию комплекса «мессенджер-рецептор», который попадает в клетку посредством опосредованного рецепторами эндоцитоза. Этот процесс увеличивает скорость деградации рецепторов внутри клетки. Следовательно, при повышенных концентрациях мессенджеров количество рецепторов плазматической мембраны постепенно уменьшается в процессе понижающей регуляции.
Также происходит обратное изменение, называемое повышающей регуляцией. Клетки, подвергающиеся длительному воздействию очень низких концентраций мессенджера, могут вырабатывать гораздо больше рецепторов для этого мессенджера, тем самым повышая чувствительность к нему. Чем больше количество рецепторов доступно для связывания лиганда, тем выше вероятность связывания. Например, при повреждении нервов, ведущих к мышце, доставка нейромедиаторов уменьшается или полностью прекращается. Со временем в таких условиях мышцы начинают сокращаться в ответ на гораздо меньшее количество нейромедиатора, чем обычно, потому что рецепторы подверглись повышающей регуляции, что привело к росту чувствительности.
Один из механизмов повышающей регуляции включает привлечение к плазматической мембране внутриклеточных везикул, содержащих многочисленные рецепторные белки в своих мембранах. Эти везикулы сливаются с плазматической мембраной, тем самым встраивая свои рецепторы в мембрану. Регуляция рецепторов в обоих направлениях (повышающая и понижающая регуляция) является прекрасным примером общего физиологического принципа гомеостаза, поскольку она способствует возвращению силы сигнала к норме, когда концентрация мессенджеров колеблется выше или ниже нормального уровня. Понимание этих механизмов лежит в основе разработки многих лекарственных препаратов, от бета-блокаторов до антигистаминных средств, и продолжает расширять возможности фармакологии и персонализированной медицины.
Сведения об авторах и источниках:
Авторы: Эрик П. Видмайер, Бостонский университет, Хершел Рафф, Медицинский колледж Висконсина, Медицинский центр Авроры Сент-Люк, Кевин Т. Странг, Университет Висконсин-Мэдисон
Источник: Физиология человека: механизмы функционирования организма
Данные публикации будут полезны студентам биологических и медицинских специальностей, начинающим специалистам в области клеточной биологии, биофизики и физиологии, а также всем, кто интересуется основами мембранного транспорта и регуляции клеточного гомеостаза.
Дата добавления: 2026-04-29; просмотров: 238;
Поиск по сайту
Узнать еще
- Аквапорины и осмос: механизмы водного транспорта и регуляции объема клеток
- Аллотропия химических элементов: Удивительные превращения формы и свойств веществ
- Архитектура компьютерных сетей: типы, оборудование и модель OSI
- Аэродинамика полета птиц: механизмы парения и поступательного движения
- Ведомственный и производственный экологический контроль: задачи и проблемы взаимодействия
- Вибрационное опыление и эволюция пчел: механизмы специализации и экологии
- Виды и механизмы биржевых сделок: от кассовых операций до фьючерсов
- Влияние лесных пожаров на биоразнообразие почв: механизмы и последствия

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
