Сигнальные пути клетки: механизмы активации рецепторов жирорастворимыми и водорастворимыми посредниками
Связывание химического мессенджера с его рецептором вызывает конформационные изменения (изменение третичной структуры) в рецепторе — событие, известное как активация рецептора. Эта активация представляет собой начальный этап, ведущий к реакции клетки на мессенджер. Клеточные реакции могут принимать форму изменений: (1) проницаемости, транспортных свойств или электрического состояния плазматической мембраны; (2) метаболизма; (3) секреторной активности; (4) скорости пролиферации и дифференцировки; или (5) сократительной или иной активности. Несмотря на разнообразие реакций, существует общий знаменатель: все реакции напрямую связаны с изменениями специфических клеточных белков.
Например, генерирование электрических сигналов в нейронах, вызванное нейромедиаторами, отражает изменённую конформацию мембранных белков (ионных каналов), через которые диффундируют ионы. Аналогичным образом, изменения скорости секреции глюкозы печенью, вызванные гормоном адреналином, отражают изменение активности и концентрации ферментов в путях синтеза глюкозы. Мышечное сокращение, вызванное нейромедиатором ацетилхолином, является результатом изменения конформации сократительных белков. Активация рецептора посредством мессенджера — это только первый шаг, ведущий к окончательному ответу клетки (сокращение, секреция и т.д.).
Различные последовательности событий, которые связывают активацию рецептора с клеточными реакциями, называются путями передачи сигнала. «Пути» обозначают специфичные для клетки механизмы, связанные с различными посредниками. Пути передачи сигнала принципиально отличаются между жирорастворимыми и водорастворимыми посредниками, поскольку их рецепторы расположены в разных местах: внутриклеточно для жирорастворимых и на плазматической мембране для водорастворимых.
Пути, инициируемые жирорастворимыми посредниками. К жирорастворимым посредникам относятся гидрофобные вещества, такие как стероидные гормоны и гормоны щитовидной железы. Их рецепторы принадлежат к большому семейству внутриклеточных рецепторов, называемых ядерными рецепторами, которые имеют сходную структуру (см. рис. 5.1, б) и механизмы действия. Хотя были идентифицированы рецепторы на плазматической мембране для некоторых из этих мессенджеров, большинство рецепторов в этом семействе являются внутриклеточными. В некоторых случаях неактивные рецепторы располагаются в цитозоле и перемещаются в ядро после связывания со своим лигандом; однако большинство неактивных рецепторов уже находятся в ядре клетки, где они связываются с соответствующими лигандами и активируются ими. В обоих случаях активация рецептора приводит к изменению скорости транскрипции одного или нескольких генов в конкретной клетке.
В наиболее распространённом случае переносчик диффундирует из капилляров из плазмы в интерстициальную жидкость (см. рис. 1.3). Оттуда мессенджер диффундирует через липидные бислои плазматической мембраны и ядерную оболочку, проникает в ядро и связывается с находящимся там рецептором (рис. 5.4). Затем активированный рецепторный комплекс функционирует в ядре в качестве фактора транскрипции — регуляторного белка, который непосредственно влияет на транскрипцию генов. Гормонально-рецепторный комплекс связывается с ДНК в регуляторной области гена, обычно увеличивая скорость транскрипции этого гена в мРНК. Молекулы мРНК выходят из ядра, чтобы направлять синтез белка, кодируемого геном, на рибосомах. Результатом является увеличение клеточной концентрации белка и/или скорости его секреции, что в конечном итоге определяет реакцию клетки на мессенджер. Например, если белок, кодируемый геном, является ферментом, ответом клетки является увеличение скорости реакции, катализируемой этим ферментом.
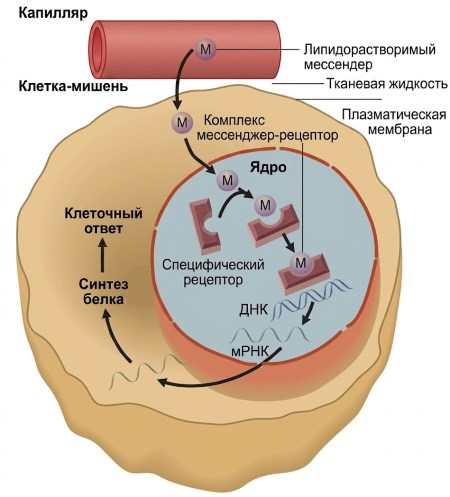
Рисунок 5.4. Механизм действия жирорастворимых посредников. На этом рисунке показан рецептор (в упрощённом виде) для этих посредников в ядре. В некоторых случаях несвязанный рецептор находится в цитозоле, а не в ядре, и в этом случае связывание происходит там, и активированный комплекс мессенджер-рецептор затем перемещается в ядро. Для простоты показано, что один мессенджер связывается с одним рецептором. Однако во многих случаях два комплекса мессенджер-рецептор должны соединиться вместе, чтобы активировать ген.
Важны ещё два момента. Во-первых, несколько генов могут контролироваться одним типом рецепторов. Например, гормон надпочечников кортизол действует через свой внутриклеточный рецептор, активируя многочисленные гены, участвующие в координированном контроле клеточного метаболизма и энергетического баланса. Во-вторых, в некоторых случаях активированный рецептор может снижать, а не усиливать транскрипцию гена или генов. Кортизол ингибирует транскрипцию нескольких генов, белковые продукты которых опосредуют воспалительные реакции после травмы или инфекции; по этой причине кортизол обладает важным противовоспалительным действием.
Пути, инициируемые водорастворимыми переносчиками. Водорастворимые переносчики не могут легко проникать в клетки путём диффузии через липидный бислой плазматической мембраны. Вместо этого они оказывают своё действие, связываясь с внеклеточной частью рецепторных белков, встроенных в плазматическую мембрану. К водорастворимым посредникам относятся большинство полипептидных гормонов, нейротрансмиттеров, а также паракринные и аутокринные соединения. Механизмы передачи сигналов, инициируемые водорастворимыми посредниками, можно разделить на четыре типа, показанные на рисунке 5.5.
Для этого обсуждения необходимы три общих термина. Во-первых, внеклеточные химические посредники (гормоны или нейромедиаторы), которые достигают клетки и связываются с её специфическими рецепторами на плазматической мембране, часто называют первыми посредниками. Вторичные посредники — это вещества, которые попадают в цитоплазму или образуются в ней в результате активации рецептора первым посредником; они распространяются по всей клетке и служат химическими ретрансляторами от плазматической мембраны к биохимическим механизмам внутри клетки. В-третьих, протеинкиназа — это фермент, который фосфорилирует другие белки, передавая им фосфатную группу из АТФ. Фосфорилирование аллостерически изменяет третичную структуру белка и, следовательно, изменяет его активность. Разные белки реагируют по-разному — некоторые активируются, некоторые инактивируются.
Существует множество различных протеинкиназ, каждая из которых способна фосфорилировать только определённые белки, и они участвуют в путях передачи сигнала, которые могут включать каскады, в которых одна протеинкиназа активирует другую, в конечном счёте фосфорилируя ключевые белки, такие как транспортеры, метаболические ферменты, ионные каналы и сократительные белки. Ферменты, которые осуществляют обратный процесс — протеинфосфатазы — дефосфорилируют белки, а также участвуют в передаче сигнала, останавливая его, как только происходит клеточная реакция.
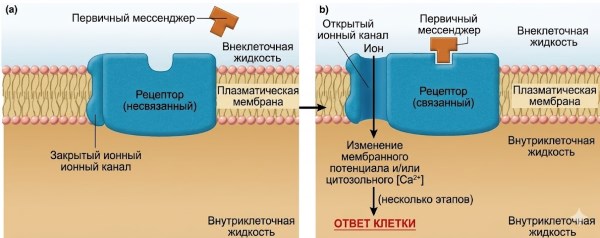
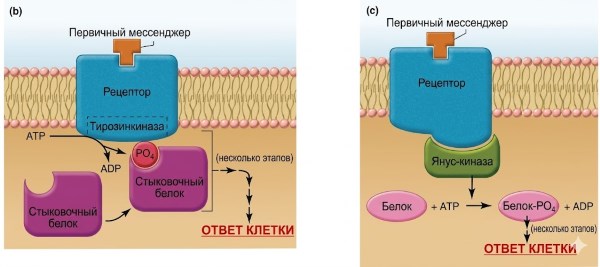
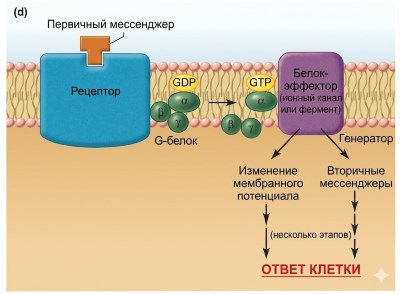
Рисунок 5.5. Механизмы действия водорастворимых посредников (на этом и последующих рисунках они обозначены как «первые посредники»). (а) Механизм передачи сигнала, в котором рецепторный комплекс включает ионный канал. Обратите внимание, что рецептор существует в двух конформациях — в несвязанном и связанном состояниях. Именно связывание первого мессенджера с его рецептором запускает конформационное изменение, которое приводит к открытию канала. Примечание: Конформационные изменения также происходят на панелях b–d, но для простоты показано только связанное состояние. (b) Механизм передачи сигнала, при котором сам рецептор функционирует как фермент, обычно тирозинкиназа. (c) Механизм передачи сигнала, при котором рецептор активирует янус-киназу в цитоплазме. (d) Механизм передачи сигнала, включающий G-белки. Когда GDP связывается с альфа-субъединицей G-белка, белок существует в виде неактивной тримерной молекулы. Связывание GTP с альфа-субъединицей вызывает диссоциацию альфа-субъединицы, которая затем активирует эффекторный белок.
Передача сигналов рецепторами — лиганд-зависимыми ионными каналами. В одном из типов рецепторов плазматической мембраны для водорастворимых мессенджеров белок, который действует как рецептор, также является ионным каналом (см. рис. 4.7). Активация рецептора первым посредником (лигандом) приводит к такому изменению конформации, что он образует открытый канал через плазматическую мембрану (рис. 5.5а). Эти каналы известны как лиганд-управляемые ионные каналы (описаны в главе 4) и особенно распространены в плазматических мембранах нейронов и скелетных мышц (главы 6 и 9). Открытие ионных каналов, управляемых лигандом, в ответ на связывание первого мессенджера увеличивает суммарную диффузию через плазматическую мембрану одного или нескольких типов ионов, специфичных для этого канала. Как было показано в главе 4 (см. рис. 4.6), такое изменение диффузии ионов приводит к изменению электрического заряда, или мембранного потенциала, клетки — это изменение мембранного потенциала является реакцией клетки на мессенджер. Кроме того, когда канал является каналом Ca²⁺, его открытие увеличивает цитозольную концентрацию Ca²⁺, что является ещё одним важным событием во многих сигнальных системах.
Передача сигналов рецепторами, функционирующими как ферменты. Другие рецепторы водорастворимых мессенджеров на плазматической мембране обладают собственной ферментативной активностью. За одним важным исключением, все рецепторы, обладающие собственной ферментативной активностью, являются протеинкиназами (рис. 5.5, б). Подавляющее большинство из них специфически фосфорилируют остатки тирозина; следовательно, эти рецепторы известны как рецепторные тирозинкиназы. Типичная последовательность такова: связывание специфического мессенджера с рецептором изменяет его конформацию таким образом, что активируется его ферментативная часть (расположенная на цитоплазматической стороне), что приводит к аутофосфорилированию рецептора (рецептор фосфорилирует некоторые из своих собственных остатков тирозина). Вновь созданные фосфотирозины в цитоплазматической части служат местами стыковки для цитоплазматических белков. Связанные стыковочные белки затем связывают и активируют другие белки, которые, в свою очередь, активируют один или несколько сигнальных путей внутри клетки. Общим знаменателем является то, что все эти пути включают активацию цитоплазматических белков путём фосфорилирования.
Существует одно физиологически важное исключение: рецептор, который функционирует и как рецептор, и как гуанилилциклаза, катализируя образование в цитоплазме циклического GMP (цГМФ). В свою очередь, цГМФ функционирует как второй посредник для активации цГМФ-зависимой протеинкиназы, которая фосфорилирует специфические белки, опосредующие реакцию клетки. Как описано в главе 7, такие рецепторы в изобилии представлены на сетчатке глаза, где они обрабатывают визуальную информацию. Этот путь используется лишь небольшим количеством посредников. Кроме того, в цитоплазме некоторых клеток присутствуют ферменты гуанилилциклазы; первый посредник — газообразный оксид азота (NO) — проникает в цитозоль и, соединяясь с гуанилилциклазой, запускает образование цГМФ. Оксид азота — это жирорастворимый газ, образующийся из аминокислоты аргинина под действием синтазы оксида азота, присутствующей во многих типах клеток, включая клетки, выстилающие кровеносные сосуды. Высвобождаясь, NO действует локально паракринным образом, расслабляя гладкую мускулатуру определённых кровеносных сосудов, что способствует расширению сосудов — важной части гомеостатического контроля артериального давления.
Передача сигналов рецепторами, взаимодействующими с цитоплазматическими янус-киназами. В этой категории механизмов передачи сигналов для водорастворимых мессенджеров (рис. 5.5, в) ферментативная активность — опять же, активность тирозинкиназы — зависит не от рецептора, а от семейства отдельных цитоплазматических киназ, называемых янус-киназами (JAK), которые связаны с рецептором. Рецептор и связанная с ним янус-киназа функционируют как единое целое. Связывание первого мессенджера с рецептором вызывает изменение конформации, приводящее к активации янус-киназы. Разные рецепторы взаимодействуют с разными представителями семейства янус-киназ, и разные янус-киназы фосфорилируют разные белки-мишени, многие из которых действуют как факторы транскрипции. Результатом работы этих путей является синтез новых белков, которые опосредуют реакцию клетки на первый мессенджер. Одним из важных примеров сигналов, передаваемых главным образом через рецепторы, связанные с янус-киназами, являются сигналы цитокинов — белков, секретируемых клетками иммунной системы, которые выполняют важнейшие функции в иммунной защите.
Передача сигналов с помощью рецепторов, связанных с G-белком. Четвёртая категория сигнальных путей для водорастворимых мессенджеров на сегодняшний день является самой многочисленной и включает сотни различных рецепторов (рис. 5.5, г). С неактивным рецептором связан белковый комплекс на цитозольной поверхности плазматической мембраны, относящийся к семейству G-белков. G-белки содержат три субъединицы: альфа, бета и гамма. Альфа-субъединица может связывать GDP и GTP; бета- и гамма-субъединицы помогают закрепить альфа-субъединицу в мембране. Связывание первого мессенджера с рецептором изменяет конформацию рецептора, повышая сродство альфа-субъединицы к GTP. При связывании с GTP альфа-субъединица отделяется от бета- и гамма-субъединиц. Эта диссоциация позволяет активированной альфа-субъединице соединиться с другим белком плазматической мембраны — либо ионным каналом, либо ферментом, — который служит в качестве эффекторного белка, опосредующего следующие шаги в последовательности, ведущей к реакции клетки.
Таким образом, G-белок служит переключателем для соединения рецептора с ионным каналом или ферментом в плазматической мембране; следовательно, эти рецепторы известны как рецепторы, связанные с G-белком. G-белок может вызывать открытие ионного канала, что приводит к изменению электрических сигналов или (в случае Ca²⁺-каналов) к изменению цитозольной концентрации Ca²⁺. Альтернативно, G-белок может активировать или ингибировать мембранный фермент, с которым он взаимодействует; такие ферменты при активации вызывают образование вторичных мессенджеров внутри клетки. Как только альфа-субъединица G-белка активирует свой эффекторный белок, активность GTPase, присущая альфа-субъединице, расщепляет GTP на GDP и Pi, делая альфа-субъединицу неактивной и позволяя ей рекомбинировать со своими бета- и гамма-субъединицами.
Существует несколько подсемейств G-белков плазматической мембраны, в каждом из которых есть несколько различных представителей, и один рецептор может быть связан с несколькими типами G-белков. Более того, некоторые G-белки могут быть связаны с несколькими типами эффекторных белков плазматической мембраны. Таким образом, рецептор, активируемый первым мессенджером, посредством своих связей с G-белками может приводить в действие различные белки плазматической мембраны, такие как ионные каналы и ферменты, которые, в свою очередь, индуцируют различные клеточные процессы. Чтобы проиллюстрировать основные моменты, касающиеся G-белков, эффекторных белков плазматической мембраны, вторичных мессенджеров и протеинкиназ, в последующих разделах описаны два наиболее распространённых эффекторных белковых фермента, регулируемых G-белками, — аденилатциклаза и фосфолипаза С.
Сведения об авторах и источниках:
Авторы: Эрик П. Видмайер, Бостонский университет, Хершел Рафф, Медицинский колледж Висконсина, Медицинский центр Авроры Сент-Люк, Кевин Т. Странг, Университет Висконсин-Мэдисон
Источник: Физиология человека: механизмы функционирования организма
Данные публикации будут полезны студентам биологических и медицинских специальностей, начинающим специалистам в области клеточной биологии, биофизики и физиологии, а также всем, кто интересуется основами мембранного транспорта и регуляции клеточного гомеостаза.
Дата добавления: 2026-04-29; просмотров: 176;
Поиск по сайту
Узнать еще
- Аквапорины и осмос: механизмы водного транспорта и регуляции объема клеток
- Атомарные запоминающие среды. Пути реализации проблемы
- Аэродинамика полета птиц: механизмы парения и поступательного движения
- Вибрационное опыление и эволюция пчел: механизмы специализации и экологии
- Виды и механизмы биржевых сделок: от кассовых операций до фьючерсов
- Влияние лесных пожаров на биоразнообразие почв: механизмы и последствия
- Внутренние и внешние источники энергии Земли: механизмы тектоники плит, мантийной конвекции и климатических циклов
- Возмещение вреда здоровью от экологических правонарушений: правовые механизмы и компенсации

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
