Аэродинамика полета птиц: механизмы парения и поступательного движения
Изучение движения птиц в воздухе исторически являлось одной из ключевых задач для естествоиспытателей, стремившихся познать природу ежедневно наблюдаемых явлений. Несмотря на кажущуюся простоту, способность пернатых с легкостью преодолевать сопротивление воздушной среды долгое время оставалась загадочной. В то время как основные принципы машущего полета в значительной степени объяснены, некоторые частные явления, такие как статическое парение, продолжают привлекать внимание исследователей. Эти феномены часто противоречат интуитивным ожиданиям, требуя для своего объяснения применения строгих физико-математических методов.
Особый интерес представляет длительное парение крупных птиц, таких как орлы или аисты, которые способны часами удерживаться в воздухе без видимых взмахов крыльями. Объяснение этого явления начинается с рассмотрения планирования. Птица с распростертыми крыльями большой площади опускается медленнее компактного объекта аналогичной массы, подобно парашюту. Сопротивление воздуха, создающее подъемную силу, прямо пропорционально площади несущей поверхности. Таким образом, планирование возможно после набора скорости активными взмахами, когда горизонтальное движение компенсирует вертикальное падение.
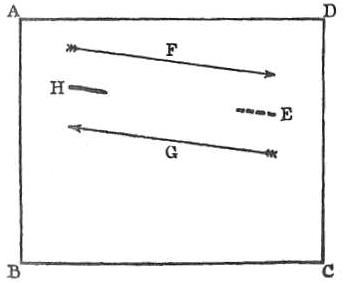
Рис. 15. Схематичное изображение птицы (H), парящей в восходящем потоке. Плоскость ABCD представляет воздушный слой, стрелка F — направление планирования в неподвижном воздухе (к точке E).
Однако стационарное парение, при котором не наблюдается ни снижения, ни поступательного скольжения, нельзя объяснить одним лишь планированием. Наблюдения показывают, что такое парение почти всегда имеет круговой или спиралевидный характер и осуществляется при наличии ветра. Птица использует динамические потоки воздуха, приводя крылья в определенное наклонное положение относительно вектора ветра. Ключевую роль играют восходящие воздушные течения (термики), часто образующиеся в нижних слоях атмосферы. Птица, попавшая в такой поток, может парить, компенсируя силу тяжести за счет энергии движущегося воздуха.
Более сложное объяснение предполагает наличие разнонаправленных воздушных течений на разных высотах. Если в верхнем слое существует устойчивый наклонно восходящий поток, близкий к горизонтальному, птица может оставаться в одной точке относительно земли. В этом случае движение воздушной массы в одном направлении полностью компенсирует аэродинамическое скольжение птицы в противоположном, что приводит к эффекту статического зависания. Точность подобных наблюдений часто осложнена большими дистанциями, на которых трудно зафиксировать минимальные изменения высоты или положения.
Другой фундаментальный вопрос аэродинамики полета касается механизма поступательного движения вперед без явного отталкивания от воздуха назад, по аналогии с гребцом. Наблюдение за медленно летящими птицами, такими как ворона или чайка, не выявляет явного движения крыльями назад. Объяснение кроется в сложной геометрии и кинематике взмаха. При опускании крыла его передняя кромка располагается ниже задней кромки, создавая угол атаки.
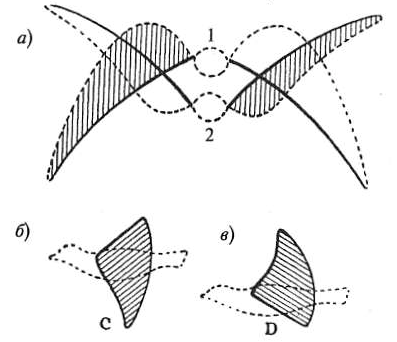
Рис. 16а. Поперечные разрезы птицы в разных фазах полета. Позиция 1: завершение удара крыла вниз (вид на верхнюю сторону). Позиция 2: завершение подъема крыла вверх (вид на нижнюю сторону). Пунктир обозначает заднюю кромку, сплошная линия — переднюю.
Рис. 16б и 16в. Вид птицы сбоку: б — верхняя сторона крыла, в — нижняя сторона. Показано, что передняя кромка расположена глубже задней.
Таким образом, при ударе крыла вниз оно взаимодействует с воздухом не строго вертикально, а отбрасывает воздушные массы назад и вниз. Согласно третьему закону Ньютона, это создает силу тяги, продвигающую птицу вперед. На схемах наклон крыла для наглядности преувеличен; в реальности достаточно минимального угла, который формируется не всей плоскостью, а главным образом гибкой задней кромкой крыла, образованной маховыми перьями.
Несущую поверхность крыла формируют маховые перья, отходящие от жесткого каркаса, состоящего из плечевой кости и костей пальцев. Задний край, лишенный жесткой опоры, обладает высокой гибкостью. При быстром ударе вниз под давлением воздуха эта часть крыла прогибается вверх, особенно сильно в области верхушки крыла, имеющей наибольшую линейную скорость. Именно это локальное изменение профиля, а не наклон всего крыла, является основной причиной генерации тяги. Экспериментально существование тяги легко проверить, совершая резкие движения в воздухе высушенным куриным крылом, при котором ощущается рывок передней кромки вперед.
Для демонстрации явления может быть использована упрощенная модель. Рама из согнутой бамбуковой удочки длиной около двух метров, стянутая тетивой, обтягивается тканью. При резком вертикальном ударе такой моделью вниз гибкий задний край, закрепленный только на шнуре, сильно загибается вверх. Аэродинамическое сопротивление создает настолько значительный вращающий момент, что удержать модель в вертикальной траектории без проворачивания вперед становится практически невозможно, что наглядно иллюстрирует возникновение силы тяги.
Механизм подъема крыльев после рабочего удара вниз также заслуживает внимания. После расслабления мышц-депрессоров, опускающих крыло, сопротивление воздуха, действующее на его нижнюю поверхность, пассивно возвращает крыло в верхнее положение. При этом тело птицы несколько опускается. Хотя у птиц существуют и активные мышцы-подниматели (элеваторы), данные исследований указывают, что в большинстве режимов полета подъем крыла является преимущественно пассивной фазой цикла. Таким образом, взмах представляет собой ритмичный процесс, где активное мощное опускание сочетается с относительно пассивным, энергосберегающим подъемом, что оптимизирует затраты энергии на полет.
Таким образом, полет птиц представляет собой результат сложной интеграции морфологических адаптаций и аэродинамических принципов. Парение обеспечивается использованием внешней энергии ветра и восходящих потоков, а поступательное движение в машущем полете — за счет сложной кинематики крыла, преобразующей вертикальные удары в горизонтальную тягу благодаря гибкости его задней кромки. Эти механизмы, отточенные миллионами лет эволюции, продолжают служить источником вдохновения для бионики и разработки современных летательных аппаратов.
Сведения об авторах и источниках:
Авторы: В. Гампсон, К. Шеффер
Источник: Парадоксы природы
Данные публикации будут полезны студентам физических и технических специальностей, изучающих механику и принципы работы простых механизмов, начинающим инженерам и конструкторам, интересующимся эргономикой и оптимизацией транспортных средств, а также всем, кто увлекается историей техники и неочевидными физическими явлениями в повседневной жизни.
Дата добавления: 2026-02-16; просмотров: 178;
Поиск по сайту
Узнать еще
- Бумеранг: аэродинамика и механика полета возвращающегося оружия
- Виды и механизмы биржевых сделок: от кассовых операций до фьючерсов
- Влияние лесных пожаров на биоразнообразие почв: механизмы и последствия
- Возмещение вреда здоровью от экологических правонарушений: правовые механизмы и компенсации
- Глобальный климат: Система, баланс и механизмы циркуляции
- Гражданские иски в экологическом праве: зарубежный опыт и механизмы защиты
- Демонстрация фотоэффекта и направленного движения зарядов в воздухе при атмосферном давлении

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
