Нервная система акулы: особенности строения и функции черепных нервов
Систематическое положение объекта. Подтип Позвоночные (Vertebrate) — представляет группу животных, обладающих развитым осевым скелетом и центральной нервной системой, защищенной черепом и позвоночным столбом. Класс Хрящевые рыбы (Chondrichthyes) включает организмов с хрящевым скелетом, которые представляют древнюю эволюционную ветвь водных позвоночных. Подкласс Пластиножаберные (Elasmobranchii) характеризуется наличием жаберных щелей без крышек и отсутствием плавательного пузыря. Отряд Катранообразные (Squaliformes) объединяет акул с колючими плавниками, а типичным представителем является Колючая акула (Squalus acanthias L.), широко используемая в учебных целях для изучения анатомии хрящевых рыб.
Материал и оборудование для исследования. Для качественного изучения нервной системы акулы на лабораторных занятиях необходимы специально подготовленные препараты и инструментарий. На одного-двух студентов требуется голова акулы, хранящаяся в 70%-ном растворе спирта с предварительно отпрепарированным головным мозгом и черепными нервами. Для работы с препаратом необходимы ванночка, пинцет, две препаровальные иглы и лупа с увеличением х4-6, позволяющая рассмотреть мелкие структуры нервной системы. Данный набор инструментов обеспечивает возможность детального препарирования и изучения анатомических особенностей мозга акулы.
Задание по изучению препарата. В процессе лабораторной работы необходимо последовательно рассмотреть все отделы головного мозга акулы и проследить отхождение черепных нервов с их наиболее крупными ветвями. Особое внимание следует уделить глазным мышцам и подходящим к ним нервным окончаниям, что позволяет понять принципы иннервации глазодвигательного аппарата. Результаты исследования оформляются в виде двух рисунков: первый демонстрирует общий вид мозга с отходящими нервами, второй — глаз с его мышцами и иннервирующими их нервами. Дополнительное задание включает знакомство со строением стато-акустического органа (внутреннего уха) по демонстрационному препарату или модели.
Общая характеристика головного мозга акулы. Головной мозг акулы представляет собой сложно организованный орган, состоящий из пяти классических отделов, характерных для всех позвоночных животных. Передний мозг (telencephalon) у акуловых рыб достигает крупных размеров и нечетко разделен на правое и левое полушария, что отражает эволюционный уровень развития хрящевых рыб. В передней части полушарий отчетливо заметны сильно развитые обонятельные доли, что свидетельствует о важной роли обоняния в жизнедеятельности акул как активных хищников. Задняя часть переднего мозга плавно переходит в промежуточный мозг (diencephalon), который на препарате визуализируется как характерное углубление между вздутиями соседних отделов.
Строение среднего и заднего отделов мозга. Средний мозг (mesencephalon) у акулы представлен крупными парными вздутиями — зрительными долями, которые своим объемом прикрывают остальную массу этого отдела, что связано с важной ролью зрения в ориентации и охоте. Мозжечок (cerebellum) у акуловых рыб достигает значительного развития и анатомически налегает спереди на средний мозг, а сзади — на продолговатый мозг, что обеспечивает высокую координацию движений при плавании. Продолговатый мозг (myelencephalon) является последним отделом головного мозга, и в его центральной части сверху отчетливо видна ромбовидная ямка (четвертый желудочек) — полость этого отдела, которая в естественном состоянии затянута сосудистой оболочкой, удаляемой при препарировании.
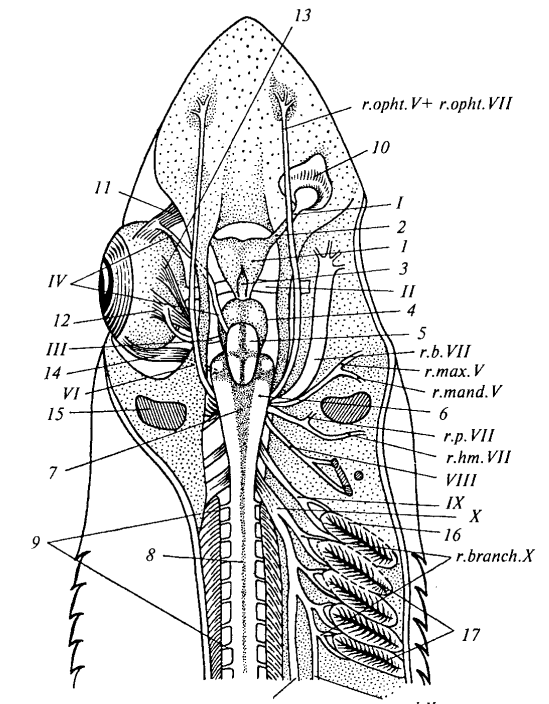
Рис. 26. Головной мозг и головные нервы колючей акулы (со спинной стороны; правый глаз удален): 1 — передний мозг; 2 — обонятельная доля переднего мозга; 3 — промежуточный мозг; 4 — средний мозг; 5 — мозжечок; 6 — продолговатый мозг; 7 — ромбовидная ямка; 8 — спинной мозг; 9 — спинномозговые нервы; 10 — обонятельный мешок; 11 — верхняя косая мышца глаза; 12 — верхняя прямая мышца глаза; 13 — внутренняя прямая мышца глаза; 14 — наружная прямая мышца глаза; 15 — брызгальце; 16 — первая жаберная щель; 17 — вторая—пятая жаберные щели; I — обонятельный тракт; II — зрительный нерв; III — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; r.opht. V — глазничная ветвь; r.max. V — верхнечелюстная ветвь; r.mand. V — нижнечелюстная ветвь; VI — отводящий нерв; VII — лицевой нерв: r.opht.VII — глазничная ветвь; r.b.VII — щечная ветвь; r.p.VII — нёбная ветвь; r.hm.VII — подъязычная ветвь; VIII — слуховой нерв; IX — языкоглоточный нерв; X — блуждающий нерв; r.branch.X — жаберные ветви; r.spl.X — внутренностная ветвь; r.lat.X — боковая ветвь
Система головных нервов акулы. Характер отхождения головных (черепных) нервов у акуловых рыб является типичным для всех позвоночных животных и служит важным систематическим признаком. От головного мозга акулы симметрично с обеих сторон отходят десять пар черепных нервов, каждая из которых имеет двойное обозначение: порядковый номер римскими цифрами и анатомическое название. Эта система иннервации обеспечивает чувствительную и двигательную связь мозга со всеми органами и тканями головы и части туловища. Изучение топографии черепных нервов имеет фундаментальное значение для понимания эволюции нервной системы позвоночных.
Обонятельный и зрительный нервы. I пара — обонятельный нерв (nervus olfactorius) у акуловых рыб, как и у других позвоночных, представлен выростами обонятельных долей переднего мозга — обонятельными трактами, которые заканчиваются расширениями у обонятельных капсул. Сам обонятельный нерв представляет собой комплекс коротких чувствующих волокон, соединяющих эпителий обонятельного мешка с обонятельной луковицей, что обеспечивает восприятие запахов в водной среде. II пара — зрительный нерв (n.opticus) фактически является зрительным трактом, отходящим от нижнебоковой поверхности промежуточного мозга и проникающим через отверстие в стенке глазницы в глазное яблоко. Этот чисто чувствующий нерв распространяется по внутренней поверхности сетчатки, передавая зрительную информацию в мозг для обработки.
Иннервация глазодвигательного аппарата. Движения глазного яблока у всех позвоночных осуществляются шестью глазными мышцами, каждая из которых одним концом прикрепляется к стенке глазницы, а другим — к определенному участку глазного яблока. От передней стенки глазницы отходят две косые мышцы: верхняя косая (m.obliquus superior) и нижняя косая (m.obliquus inferior), обеспечивающие вращательные движения глаза. От задней стенки глазницы пучком отходят четыре прямые мышцы: нижняя прямая (m.rectus inferior), внутренняя прямая (m.rectus internus), верхняя прямая (m.rectus superior) и наружная прямая (m.rectus externus). Эти мышцы иннервируются тремя парами головных нервов — III, IV и VI, которые являются чисто двигательными и обеспечивают точную координацию движений глаз.
Глазодвигательный и блоковый нервы. III пара — глазодвигательный нерв (n.oculomotorius) отходит от дна среднего мозга и после прохождения через стенку черепа делится у основания прямых мышц на ветви к четырем мышцам. Этот нерв иннервирует нижнюю косую, а также нижнюю, внутреннюю и верхнюю прямые мышцы, обеспечивая основные движения глазного яблока. IV пара — блоковый нерв (n.trochlearis) представляет собой тонкую нить, отходящую от задневерхней части среднего мозга и выходящую из-под мозжечка, что хорошо видно на препарате. Пройдя через переднюю часть стенки глазницы, этот нерв разветвляется исключительно в верхней косой мышце глаза, обеспечивая ее специфическую функцию.
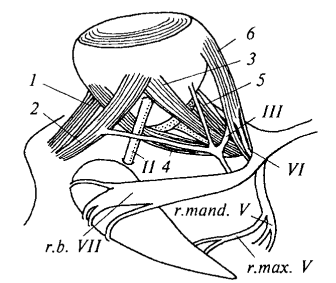
Рис. 27. Глазные мышцы акулы и иннервирующие их нервы на нижней стороне левого глаза (глаз отогнут вверх): 1 — верхняя косая мышца глаза; 2 — нижняя косая мышца; 3 — нижняя прямая мышца глаза; 4 — внутренняя прямая мышца; 5 — верхняя прямая мышца; 6 — наружная прямая мышца; II — зрительный нерв; III — глазодвигательный нерв; r.mand.V — нижнечелюстная ветвь тройничного нерва; r.max.V — верхнечелюстная ветвь тройничного нерва; VI — отводящий нерв; r.b.VII — щечная ветвь лицевого нерва
Отводящий и тройничный нервы. VI пара — отводящий нерв (n.abducens) отходит от дна продолговатого мозга и сразу же уходит в дно черепной коробки, поэтому на препарате его отхождение обычно не визуализируется. Этот нерв иннервирует наружную прямую мышцу глаза, и на ее внутренней поверхности хорошо заметны его разветвления, обеспечивающие отведение глаза в латеральном направлении. V пара — тройничный нерв (n.trigeminus) отходит толстым корнем от переднебоковой поверхности продолговатого мозга и сразу делится на три крупные ветви: глазничную, верхнечелюстную и нижнечелюстную. По функции тройничный нерв является смешанным: глазничная и верхнечелюстная ветви несут чувствительные волокна, а нижнечелюстная содержит как чувствительные, так и двигательные компоненты.
Ветви тройничного нерва. Глазничная ветвь (ramus ophthalmicus) тройничного нерва проходит через глазницу вместе с одноименной ветвью лицевого нерва и иннервирует кожные органы чувств в передней части рыла. Верхнечелюстная ветвь (r.maxillaris) и нижнечелюстная ветвь (r.mandibularis) начинаются общим стволом и обособляются позади глазного яблока, обеспечивая иннервацию мышц челюстной дуги. Эти ветви также снабжают нервами зубы и кожу головы, что критически важно для хищного образа жизни акулы, поскольку обеспечивает чувствительность челюстного аппарата при захвате добычи. Сложное строение тройничного нерва отражает его ключевую роль в обеспечении как двигательной активности челюстей, так и сенсорного восприятия переднего отдела головы.
Лицевой нерв и его ветви. VII пара — лицевой нерв (n.facialis) отходит от продолговатого мозга сразу за тройничным нервом, и на препарате их основания трудно четко разграничить, что анатомически закономерно. Этот сложный нерв распадается на несколько крупных ветвей, среди которых наиболее значимыми являются подъязычная, нёбная, глазничная и щечная ветви. Подъязычная ветвь (r.hyomandibularis) проходит позади брызгальца и иннервирует подъязычную висцеральную дугу с ее мускулатурой и слизистой оболочкой. Нёбная ветвь (r.palatinus) отходит вперед от основания предыдущей и ветвится в слизистой оболочке крыши ротовой полости, обеспечивая вкусовую чувствительность.
Сенсорные ветви лицевого нерва. Глазничная ветвь (r.ophtalmicus) лицевого нерва следует вместе с одноименной ветвью тройничного нерва, иннервируя кожные органы боковой линии на спинной поверхности головы, включая специализированные электрорецепторы. Щечная ветвь (r.buccalis) аналогичным образом обеспечивает иннервацию брюшной поверхности головы, отходя крупным стволом позади глазничной ветви и проходя по дну глазницы в виде широкого плоского тяжа. Лицевой нерв, подобно тройничному, является смешанным, что обеспечивает как двигательную активность висцерального аппарата, так и чувствительное восприятие поверхности головы, включая регистрацию электрических полей, создаваемых добычей.
Слуховой и языкоглоточный нервы. VIII пара — слуховой нерв (n.acusticus) отходит от боковой поверхности продолговатого мозга и практически сразу входит в стенку черепной коробки, направляясь к внутреннему уху. Этот чисто чувствующий нерв обеспечивает передачу слуховой и вестибулярной информации от перепончатого лабиринта в центральную нервную систему для обработки и координации движений. IX пара — языкоглоточный нерв (n.glossopharyngeus) подходит к первой жаберной щели, где делится на две ветви, иннервирующие ее переднюю и заднюю поверхности. Задняя ветвь этого смешанного нерва снабжает двигательными и чувствительными волокнами всю мускулатуру первой жаберной дуги, участвуя в регуляции дыхательных движений.
Блуждающий нерв — главный парасимпатический нерв. X пара — блуждающий нерв (n.vagus) отходит от заднебоковой поверхности продолговатого мозга несколькими корешками, которые почти сразу сливаются в мощный нервный тяж, обеспечивающий иннервацию обширной области тела. В отличие от других головных нервов, блуждающий имеет обширную зону иннервации и делится на три основные ветви: жаберные, внутренностную и боковую. Четыре жаберные ветви (r.branchialis) иннервируют жаберные щели со второй по пятую, причем каждая ветвь, подходя к жаберной щели, разделяется на передне- и заднежаберную, обеспечивая как чувствительную, так и двигательную функции.
Ветви блуждающего нерва. Внутренностная ветвь (r.splanchnicus) является прямым продолжением основного ствола блуждающего нерва и проникает в брюшную полость, где иннервирует внутренние органы — сердце, пищеварительный тракт и другие висцеральные структуры. Боковая ветвь (r.lateralis) располагается более поверхностно и обеспечивает иннервацию органов боковой линии туловища и хвоста, что позволяет акуле воспринимать малейшие колебания воды. Блуждающий нерв является смешанным, содержащим как чувствительные, так и двигательные волокна, и представляет собой эволюционно древнюю структуру, обеспечивающую парасимпатическую иннервацию внутренних органов. У высших позвоночных обособляются еще две пары головных нервов: XI пара — добавочный нерв (n.accessorius) и XII пара — подъязычный нерв (n.hypoglossus).
Спинномозговые нервы. Спинномозговые нервы (n.spinalis) отходят посегментно от спинного мозга (medulla spinalis) и хорошо видны на препарате вдоль всего позвоночного столба. Каждый такой нерв иннервирует соответствующий сегмент тела и по своей функции является смешанным, включая чувствительные волокна от кожи и двигательные волокна к мускулатуре. Метamerическое расположение спинномозговых нервов отражает исходную сегментарность тела позвоночных и обеспечивает точную локальную регуляцию движений. Взаимодействие головных и спинномозговых нервов создает целостную систему иннервации всего организма акулы.
Строение внутреннего уха (стато-акустического органа). Внутреннее ухо хрящевых рыб представлено перепончатым лабиринтом, заключенным в хрящевую слуховую капсулу, встроенную в мозговой череп позади глазницы. Три изогнутые трубочки — полукружные каналы — расположены в трех взаимно перпендикулярных плоскостях и открываются концами в преддверие (перепончатый мешок). Этот мешок подразделяется на две части: верхнюю — овальный мешочек и нижнюю — круглый мешочек, от которого отходит небольшой выступ — лагена. Полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики — отоконии, а в круглом мешочке находятся более крупные известковые образования — отолиты.
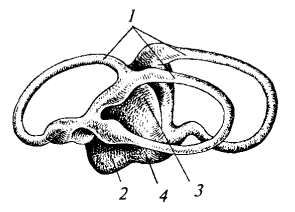
Рис. 28. Схема перепончатого лабиринта акулы: 1 — полукружные каналы; 2 — круглый мешочек; 3 — овальный мешочек; 4 — лагена
Функциональное значение внутреннего уха. Окончания слухового нерва подходят к специализированным участкам перепончатого лабиринта, покрытым чувствующим эпителием — слуховым пятнам, гребням и сосочкам. Перепончатый лабиринт воспринимает линейные и угловые ускорения, а также вибрации водной среды, обеспечивая пространственную ориентацию и равновесие. Полукружные каналы регистрируют вращательные движения головы и тела, а отолитовый аппарат реагирует на гравитацию и прямолинейные ускорения. Такое сложное строение стато-акустической системы необходимо для активного хищного образа жизни акулы в трехмерной водной среде.
Эволюционное значение хрящевых рыб. Хрящевые рыбы представляют важный этап эволюции позвоночных животных, демонстрируя более высокий уровень организации по сравнению с круглоротыми и более активные формы жизнедеятельности. Принципиальное отличие этого класса (и всех челюстноротых — Gnathostomata) от бесчелюстных (Agnatha) заключается в строении висцерального отдела черепа, состоящего из нескольких пар расчлененных подвижных дуг. Наличие челюстной дуги (обеспечивающей хватательную функцию), подъязычной дуги (обеспечивающей подвижное прикрепление челюстного аппарата) и нескольких жаберных дуг (обеспечивающих опору дыхательного аппарата) позволило интенсифицировать дыхание и облегчить захват добычи. Такие анатомические преобразования способствовали повышению активности питания и, как следствие, общему увеличению уровня обмена веществ.
Прогрессивные черты организации. У хрящевых рыб сформировался ряд прогрессивных особенностей строения по сравнению с круглоротыми, обеспечивающих их эволюционный успех. Появление парных конечностей значительно увеличило устойчивость и маневренность при плавании, что особенно важно для активных хищников. Замена хорды хрящевыми позвонками усилила опорную функцию осевого скелета при сохранении его гибкости, что способствовало активизации передвижения, где основным локомоторным органом стал хвостовой отдел. Относительные размеры головного мозга резко возросли, причем все его отделы хорошо развиты, а органы чувств — глаза, органы обоняния, слуха и сейсмосенсорная система боковой линии — достигли значительно более высокого уровня развития. Черепная коробка полностью закрывает головной мозг, а капсулы органов чувств срастаются со стенками мозговой коробки, образуя монолитный осевой череп.
Особенности дыхательной и пищеварительной систем. Челюстноротым, включая хрящевых рыб, свойствен принципиально иной тип строения органов дыхания по сравнению с круглоротыми. Эктодермальные жабры прикрепляются к подвижным жаберным дугам и состоят из многочисленных тонких жаберных лепестков, что значительно увеличивает дыхательную поверхность. Пищеварительный тракт приобретает более отчетливую дифференцировку на отделы, а поверхность всасывания увеличивается благодаря наличию в толстой кишке хорошо развитого спирального клапана. Мощное развитие пищеварительных желез — печени и поджелудочной железы — обеспечивает выработку разнообразных пищеварительных ферментов, повышая скорость и эффективность пищеварения. Вся совокупность этих прогрессивных изменений обеспечивает более высокий уровень обмена веществ и общую активизацию жизнедеятельности.
Экологические группы пластиножаберных. В подклассе Пластиножаберные (Elasmobranchii) по характеру образа жизни отчетливо выделяются две основные группы: акулы и скаты, демонстрирующие различную адаптивную эволюцию. Акулы в большинстве своем являются подвижными хищниками, обитающими в толще воды и отличающимися высокой скоростью и маневренностью плавания. Для них характерно обтекаемое торпедообразное тело, мощный хвостовой стебель с большим хвостовым плавником и крупные острые зубы, способные отрезать фрагменты от жертвы. Скаты (большинство видов) ведут придонный образ жизни и питаются малоподвижными донными животными, в связи с чем у них сформировалась уплощенная форма тела и покровительственная окраска спины. Зубы скатов уплощены и лишены острых вершин, образуя своеобразную «терку» для раздавливания раковин моллюсков и панцирей ракообразных.
Примитивные черты организации. Несмотря на прогрессивные черты, по сравнению с эволюционно более молодыми группами костных рыб многие особенности строения хрящевых рыб выглядят примитивными. Массивный хрящевой скелет увеличивает общую массу тела, а пояса парных плавников лежат в толще мускулатуры и не имеют жесткого укрепления на осевом скелете. Массивные хрящевые челюсти способны лишь к однообразным простым хватательным движениям, а адаптивные изменения, связанные с различным характером питания, ограничиваются преимущественно различиями в строении зубов. Эти ограничения не позволяют хрящевым рыбам достичь такого разнообразия жизненных форм, как у костных рыб, и обусловливают их преимущественно морское распространение.
Сравнение с костными рыбами. По сравнению с костными рыбами и их высшими представителями — костистыми рыбами, хрящевые рыбы демонстрируют более низкий уровень обмена веществ, что определяется рядом морфофизиологических особенностей. Дыхательный аппарат имеет довольно простое строение (отсутствие жаберной крышки), кишечник менее дифференцирован и короче, скорость кровотока и кислородная емкость крови ниже. Эти особенности находят выражение в ограниченном распространении хрящевых рыб (только морские обитатели) и малом разнообразии жизненных форм при ограниченном числе видов (около 600 против почти 20 тысяч видов костных рыб). Тем не менее, в морских экосистемах хрящевые рыбы остаются вполне процветающей группой, успешно конкурирующей с костными рыбами благодаря ряду уникальных адаптаций.
Уникальные адаптации хрящевых рыб. Хрящевые рыбы обладают рядом своеобразных особенностей, обеспечивающих их успешное существование в морских экосистемах. Для них характерна большая живучесть по сравнению с костными рыбами, проявляющаяся в способности к длительному голоданию, высокой устойчивости к повреждениям и значительной мощи мускулатуры. В экспериментах акула сохраняет нормальную скорость движения при дополнительной нагрузке до 25% массы тела, тогда как костистые рыбы теряют способность к движению при нагрузке 5-10%. Эти особенности связаны с относительно крупными размерами головного мозга и его высокой дифференцировкой, а также с наличием в мышцах значительного количества дыхательного пигмента — миоглобина.
Особенности размножения и осморегуляции. Специфические черты строения половой системы хрящевых рыб обеспечивают возможность внутреннего оплодотворения, что является прогрессивным признаком. Относительно крупные яйца с большим запасом питательных веществ и прочной рогоподобной скорлупой значительно снижают эмбриональную смертность, а свойственное многим видам яйцеживорождение еще более повышает эффективность размножения. Уникален у хрящевых рыб механизм осморегуляции: осмотическое давление в тканях и крови примерно равно таковому в морской воде благодаря высокому содержанию мочевины. При изменениях солености среды концентрация мочевины регулируется, приводя в соответствие осмотическое давление жидкостей тела и среды при неизменной концентрации биологически важных солей, что является уникальной адаптацией.
Заключение: эволюционный статус хрящевых рыб. Хрящевые рыбы представляют собой наиболее древнюю группу рыб, для которой характерны многие примитивные черты организации, унаследованные от далеких предков. Они сохранились до наших дней благодаря выработке своеобразных адаптивных черт организации, помогающих успешно выдерживать конкуренцию с костными рыбами — эволюционно более молодой и прогрессивной группой. Хрящевые и костные рыбы представляют собой два самостоятельных параллельных пути развития водных позвоночных животных, каждый из которых имеет свои преимущества и ограничения. Изучение нервной системы акулы позволяет понять фундаментальные принципы организации позвоночных и проследить основные направления эволюционных преобразований в этой группе животных.
Рекомендуемая литература: Гуртовой Н. Н. Систематика и анатомия хордовых животных: Краткий курс. М., 2004.
Гуртовой Н. Н., Матвеев Б. С., Дзержинский Ф. Я. Практическая зоотомия позвоночных. Низшие хордовые, бесчелюстные, рыбы. М., 1976.
Дзержинский Ф. Я. Сравнительная анатомия позвоночных животных: В 4 т. М., 1998.
Жизнь животных: В 7 т. Т. 4. Рыбы. М., 1983.
Никольский Г. В. Частная ихтиология. М., 1971.
Никольский Г. В. Экология рыб. М., 1974.
Ромер А., Парсонс Г. Анатомия позвоночных: В 2 т. М., 1992.
Суворов Е. К. Основы ихтиологии. М., 1948.
Сведения об авторах и источниках:
Авторы: Карташев Н. Н., Соколов В. Е., Шилов И. А.
Источник: Практикум по зоологии позвоночных
Данные публикации будут полезны студентам биологических специальностей, изучающих зоологию и сравнительную анатомию, а также всем, кто интересуется эволюционной морфологией и строением примитивных хордовых животных.
Дата добавления: 2026-03-05; просмотров: 145;
Поиск по сайту
Узнать еще
- ChatGPT для преподавателей: как использовать функции ИИ в обучении (практическое применение, преимущества, ограничения)
- Автоматизация проектирования и система автоматизированного проектирования ПР
- Автоматизированная система наблюдения за состоянием кардиологических больных
- Активные и пассивные операции Сберегательного банка России: структура и особенности
- Амортизационная система шасси
- Анатомические и физиологические особенности птиц. Перьевой покров
- Анатомия акулы: подробное руководство по вскрытию и систематике
- Анатомия земноводных: методика вскрытия и особенности внутреннего строения на примере травяной лягушки (Rana temporaria L.)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
