Влияние рельефа нарост и устойчивость древесных растений
Принято считать, что рельеф непосредственного влияния на жизнь растений не оказывает, но он дифференцирует поверхность и этим изменяет напряженность некоторых факторов. То есть за счет рельефа увеличивается разнообразие местообитаний и тем самым осуществляется его косвенное влияние на рост и развитие растений (А. А. Уранов, 1974; И. С. Мелехов, 1980; А. С. Тихонов, Н. М. Набатов, 1995).
Различают три категории рельефа: макрорельеф, мезорельеф и микрорельеф. Внутри той или иной категории рельефа изменение климатических и эдафических факторов зависит от высоты над уровнем моря или высоты расположения составляющих его элементов, экспозиции и крутизны склонов и других орографических особенностей.
Макрорельеф связан с горообразованием и характерен для горных стран. С высотой в горах в первую очередь меняется температура. Каждым 100 метрам подъема по вертикали соответствует снижение годовой температуры на 0,5—0,6 °С в условиях влажного воздуха и на 1 °С в условиях сухого. В связи с этим меняется режим осадков и освещения. Изменение растительности с поднятием в горы (вертикальная поясность) со времен А. Гумбольдта считается сходным с изменением растительности на равнине по мере движения в сторону полюса того же полушария (горизонтальная зональность).
Горная местность всегда представляет большое разнообразие топологических элементов, каждый со своей экспозицией, со своими углами наклона, с различной силой размывающего действия водных потоков и с почвами разной степени сформированности на тех или иных породах. Отсюда в горах нередко встречается огромное разнообразие видов, экотипов и форм растений, обусловленное пестротой экологических условий.
Световой, тепловой и водный режимы склонов связаны с экспозицией, что отражается на составе древесных пород и их продуктивности. Сочетание северных склонов с избыточно увлажненными почвами создает наиболее сильное понижение температуры почвы, а южные склоны с песчаными почвами, наоборот, способствуют повышению температуры в почве.
В связи с этим, как отмечает И. С. Мелехов (1980), в Восточной Сибири на северных склонах в лесах преобладает лиственница, на южных — сосна. На Кавказе дубовые леса чаще приурочены к южным, буковые — к северным склонам. Еловые леса (представленные разными видами) на Кавказе, в Казахстане, в горах Средней Азии обычно растут на северных, иногда на западных и восточных, но отсутствуют на южных склонах.
Мезорельеф, или рельеф средних форм в равнинных условиях, в малом масштабе повторяет картину, наблюдаемую в горах. Всхолмления на равнинах несколько “осеверяют” условия среды. Так, в равнинных районах юга европейской части России лесорастительные условия северных и северозападных склонов более благоприятны для выращивания леса, чем южных или юго-восточных, на которых успешно может расти очень ограниченное число пород (сосны обыкновенная и крымская, дуб и некоторые другие).
Мезоизменения в рельефе отражаются на снегонакоплении, содержании влаги в почве, на ее промачиваемости. Это, в свою очередь, сказывается на жизнеспособности насаждений, особенно в засушливые этапы вегетационного периода.
Микрорельеф — небольшие (на несколько десятков сантиметров) повышения или понижения поверхности почвы. Эти микропонижения или микроповышения играют существенную роль в расселении древесной растительности на равнинах. Они влияют на тепловой режим, перераспределение влаги на поверхности почвы и, как следствие, — на естественное возобновление леса.
Хорошо известно произрастание самосева и подроста ели по микроповышениям на влажных, тяжелых по механическому составу почвах, приуроченных к пониженным местоположениям мезорельефа, где эта порода находит спасение от выжимания морозом и вымокания.
С другой стороны, небольшие блюдцеобразные микропонижения в степных условиях способствуют увлажнению и рассолонцеванию почв и появлению осиновых колков, или небольших рощ. Чем ближе к районам, где летом бывает значительная засуха, тем большее значение приобретают эти микропонижения.
Таким образом, рельеф создает многообразие условий местопроизрастания и комплексов растений в разных географических областях, уменьшая или увеличивая фактическую площадь для произрастания растений.
7.5. Взаимодействие биотических факторов среды и древесных растений
Биотические факторы включают все организмы растительной и животной природы, а также микроорганизмы, которые могут оказать то или иное влияние на жизнь растений данного вида. Биотические факторы весьма разнообразны благодаря огромному видовому разнообразию организмов и специфичности жизнедеятельности каждого вида.
Биотические воздействия на растения бывают прямыми (например, поедание животными) и непрямыми (изменение среды, в которой живет растение).
Большую роль в жизни растений играет животный мир, и особенно животный мир почвы. Животные подгрызают корни, размельчают и переваривают остатки растений, передвигаются в почве, разрыхляя ее, пропускают через кишечник частицы почвы, изменяя их химический состав и строение. Влияние одних растений на другие также разнообразно. Но особенно заметное влияние на растения оказывают грибы и микроорганизмы.
Различают следующие виды взаимоотношений между организмами: хищничество, паразитизм, конкуренция, комменсализм, мутуализм (симбиоз) и др. О некоторых из них (наличие микоризы как пример симбиоза) говорилось выше.
Хищничество заключается в ловле и поедании представителями одного вида особей другого. По отношению к растениям растительноядные животные должны считаться хищниками или, по крайней мере, паразитами (Р. Риклефс, 1979; А. С. Тихонов, Н. М. Набатов, 1995).
Паразитизм (от греч. паразитос — нахлебник, от пара... — возле, мимо, вне и зитос — хлеб, пища) — это способ существования одного вида (паразита) за счет другого (хозяина). Это взаимоотношения антагонистического характера, которые необязательно ведут к гибели используемого организма, что и наблюдается в отношениях между растениями и животными (например, при паразитировании насекомых-вредителей), животными и микроорганизмами. Известен паразитизм и между деревьями (примером может служить сандаловое дерево Юго-Восточной Азии, растущее на корнях других деревьев).
Конкуренция (от лат. сопсигго — сбегаюсь, сталкиваюсь) — взаимоотношения между организмами одного вида (внутривидовая конкуренция) или разных видов (межвидовая конкуренция), соревнующимися за одни и те же ресурсы внешней среды при недостатке последних. Среди древесных растений это выливается в борьбу за свет, влагу, питательные вещества и другие ресурсы.
Комменсализм (от лат. сот — с, вместе и mensa — стол, трапеза), сотрапезничество, — форма симбиоза, при которой один из партнеров (комменсал) возлагает на другого (хозяина) регуляцию своих отношений с внешней средой, но не вступает с ним в тесные отношения. Метаболическое взаимодействие и антагонизм между партнерами в такой системе отсутствуют. Основой для комменсальных отношений могут быть общее пространство, субстрат, пища и др. Примером комменсализма служит жизнь эпифитов на стволах деревьев, которые первые используют только как место прикрепления.
Мутуализм (от лат. mutuus — взаимный) — форма симбиоза, при которой два различных организма возлагают друг на друга регуляцию своих отношений с внешней средой. При этом отношения между партнерами характеризуются взаимовыгодностью и ни один из них не может существовать без другого. Примером мутуализма может служить симбиоз древесных пород и грибов-микоризообразователей.
К числу прямых влияний растений на растения можно отнести и аллелопатию (от греч. аллелон — взаимно и патос — страдание, испытываемое воздействие) — взаимодействие растений посредством выделения биологически активных веществ (фитонцидов, колинов, антибиотиков и др.) во внешнюю среду. В одних случаях это влияние может быть стимулирующим, в других — задерживающим и даже угнетающим, в третьих — индифферентным. При этом различают: а) влияние высших растений на высшие при посредстве тормозящих веществ — колинов, б) высших растений на микроорганизмы при посредстве фитонцидов — веществ, убивающих или подавляющих рост и развитие других организмов; в) микроорганизмов на высшие растения при посредстве маразминов — веществ завяда-ния; г) микроорганизмов на микроорганизмы при посредстве антибиотиков (В. Г. Хржановский,1976).
7.6. Оценка взаимодействия “генотип — среда”
Об изменчивости, обусловленной взаимодействием “генотип — среда”, кратко упоминалось ранее (глава ). По Д. С. Фолконеру (1984), существует несколько форм, которые это взаимодействие может принимать. Так, определенное различие условий среды может иметь большее влияние на какой-нибудь один генотип, чем на все другие, или же, например, порядок ранжирования некоторых генотипов по их достоинству может изменяться, когда эти генотипы оцениваются применительно к различным условиям среды: как отмечалось ранее, генотип А может превосходить генотип В в среде X, но уступать ему в среде Y. Когда между генотипами и условиями среды есть зависимость, фенотипическая оценка особи будет не просто
Р = G + Е, как в уравнении (10.2), а она будет включать также и компоненту взаимодействия:
P = G+E+VGE . (1)
Как уже отмечалось, компонента взаимодействия дает начало дополнительному источнику изменчивости, и уравнение (6.1) преобразуется:
Vp =VG + VE+VGE (2)
Разными авторами предложен целый ряд количественных методов оценки взаимодействия “генотип — среда” для сортов растений:
G.Wricke, 1965; S. A. Eberhart, W. A. Russel, 1966; А. А. Жученко, В. С. Нестеров и др., 1980 (цит. по А. А. Жученко,1980); В. М. Роне, 1980.
Возможность применения одного из этих методов, а именно С. А. Эберхарта и В. А. Рассела, изучена на примере сортов тополя (А. П. Царев, 1985). Названная модель может быть представлена следующей зависимостью:
YiJ= Mi + 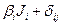 (3)
(3)
где YiJ —средняя величина параметра i-го сорта в J-м условии произрастания (i= 1, 2,..., v,j = 1,2,..., n), Mi— средняя величина признака /-го сорта во всех условиях; /5, — коэффициент регрессии, который оценивает реакцию i-го сорта на изменение условий среды (первый параметр стабильности); 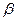 i— отклонение от линии регрессии i-го сорта при j-м условии;
i— отклонение от линии регрессии i-го сорта при j-м условии;
Jj— индекс условий среды (ИУС), полученный как среднее арифметическое всех сортов в j-м условии минус общая средняя, т. е.
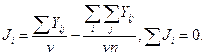 (4)
(4)
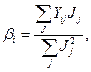 (5)
(5)
Вторым параметром стабильности служит варианса стабильности, выраженная отклонением конкретных дат от линии регрессии. Для данного случая она имеет следующее выражение:
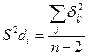 (6)
(6)
Здесь 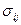 . получена как разница между теоретическими и фактическими показателями роста, т. е.
. получена как разница между теоретическими и фактическими показателями роста, т. е.
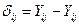 (7)
(7)
В свою очередь, теоретический рост рассчитывается по следующей формуле:
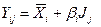 (8)
(8)
где Yij —теоретическая величина роста каждого сорта в определенных условиях среды; 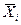 — оценка M i
— оценка M i
Для повышения объективности оценки, рассчитанной по этому методу, опыты необходимо закладывать в нескольких повторностях.
В качестве иллюстрации применения данной зависимости приведем пример оценки экологической стабильности роста разных сортов тополя в высоту на заложенном автором орошаемом сортоиспытательном участке Астраханской ЛОС в условиях светло-бурых полупустынных почв на площади более 10 гектаров, где испытывалось 11 сортов и клонов тополя.
Обобщенный анализ многолетних данных о приживаемости, сохранности, сезонном развитии, росте, устойчивости к засухе, энтомовредителям и болезням, а также о технических свойствах древесины и целлюлозного полуфабриката позволил рекомендовать для разведения в орошаемых условиях полупустыни на почвенных комплексах бэровских бугров ряд испытанных сортов тополя, в первую очередь из евроамериканских гибридов, часть из которых была использована для оценки экологической стабильности (табл.).
Дата добавления: 2017-09-01; просмотров: 4231;
Поиск по сайту
Узнать еще
- IV. Влияние катализатора на скорость реакции
- IС класс — блокаторы натриевых каналов, имеющие различное влияние на ЭРП
- Абсолютная устойчивость нелинейных систем в случае, когда W(p) не является рациональной функцией.
- Автомобили и Их Влияние на Чувство Территории
- Агрегативная устойчивость растворов ВМС и ее нарушение
- Адаптация растений и животных к световому режиму.
- Алкоголь и его влияние на физическое и психическое здоровье человека. Профилактика
- Анатомическое строение стебля однодольных и двудольных растений. Строение стебля двудольных и голосеменных древесных растений. Структура древесины

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
