Запасные вещества прокариот
| Запасное вещество | Структурные характеристики | Химический состав | Функции | Распространение |
| Гранулы гликогена | Æ20–100 нм | высокомолекулярные полимеры глюкозы | источник углерода и энергии | широко распространены |
| Гранулы поли- β-оксимасляной кислоты | Æ100–1000 нм; окружены однослойной белковой мембраной толщиной 2–3 нм | 98% полимера поли-β-оксимасляной кислоты, 2% белка | источник углерода и энергии | широко распространены |
| Гранулы полифосфата | Æ» 500 нм, зависит от условий культивирования | линейные полимеры ортофосфата | источник фосфора и энергии | широко распространены |
| Гранулы серы | Æ100–800 нм; окружены однослойной белковой мембраной толщиной 2–3 нм | включения жидкой серы | источник энергии | бактерии, окисляющие H2S |
| Углеводородные гранулы | Æ200–300 нм; окружены однослойной белковой мембраной толщиной 2–4 нм | углеводороды того же типа, что и в среде | источник углерода и энергии | представители родов Mycobacterium, Nocardia и другие, использующие углеводороды |
Функции включений:
1. Трофическая: запасные вещества служат источником питательных веществ. Так, волютиновые гранулы коринебактерий дифтерии играют роль фосфатных депо, за счет которых клетка может при недостатке фосфора в среде осуществить еще несколько делений.
2. Энергетическая: запасные вещества служат источником энергии. Поэтому при отсутствии внешних источников энергии они могут продлить время существования клетки, а у спророобразующих видов — создать условия для образования спор.
3. Дифференциально-диагностическое значение у некоторых бактерий. Напр., зерна волютина обнаруживаются у возбудителя дифтерии (C. diphtheriae) и дифтероидов(C. xerosis, вызывающего конъюнктивиты, и C. ulcerans, вызывающего дифтериеподобные поражения кожи), но отсутствуют у непатогенных ложнодифтерийных палочек(C. pseudodiphtheriticum).
Выявление включений (зерен волютина):
Волютин проявляет хромофильность и метахромазию (имея щелочную реакцию, вопринимает кислые красители), легко выявляется в клетках в виде крупных гранул, особенно при специальных методах окраски. ЦП бактериальной клетки, обладая кислой реакцией, воспринимает щелочные красители.
| 1. Окраска по Леффлеру: – на фиксированный препарат наливают щелочной раствор метиленовой синьки на 3–5 мин; – промывают водой, высушивают, наносят иммерсионное масло, микроскопируют: палочки (протоплазма) окрашиваются в светло-голубой цвет, зерна волютина (гранулы полифосфата) — в темно-синий (рис. 28). |
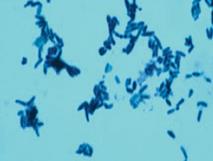 Рис.28. Окраска по Леффлеру
Рис.28. Окраска по Леффлеру
|
| 2. Окраска по Нейссеру: – на фиксированный препарат наливают уксусно-кислую метиленовую синьку на 1–2 минуты; краситель сливают, препарат промывают водой; – наносят раствор Люголя на 30–60 сек (иногда); – промывают водой; – докрашивают хризоидином 10–15 сек или везувином 30сек–3мин; – промывают водой, высушивают, микроскопируют: зерна волютина выявляются у дифтерийной палочки в виде интенсивно прокрашивающихся в темно-синий (без раствора Люголя) или черный (с раствором Люголя) цвет полюсов клетки; ЦП клетки окрашивается в нежно-желтый цвет (рис. 29). |  Рис. 29. Окраска по Нейссеру
Рис. 29. Окраска по Нейссеру
|
3. Окраска волютина флюорохромом корифосфином: в люминесцентном микроскопе бактерии желто-зеленые, зерна волютина —оранжево-красные.
ЭНДОСПОРЫ
Строение. Эндоспора —метаболически неактивная(анабиотическая, покоящаяся) форма бактерий. Эндоспоры образуются при попадании вегетативной клетки в неблагоприятную внешнюю среду.
Факторы, стимулирующие спорообразование:
– недостаток питательных веществ (прежде всего углерода и азота);
– высушивание;
– накопление в среде в большом количестве токсичных продуктов обмена веществ бактерий;
– минеральные соли, содержащие ионы кальция, магния, калия, марганца, железа, цинка, меди;
– минимальные концентрации некоторых редких металлов (кобальта, лития, кадмия, никеля);
– парааминобензойная и фолиевая кислоты.
Способностью к спорообразованию обладает небольшое число как патогенных бактерий (B. anthracis, C. perfringens, C. tetani, C. botulinum), так и непатогенных бактерий (сапрофитные представители рода Bacillus и некоторые кокки) (рис. 30). Спорообразование — непостоянный признак: у бацилл сибирской язвы споры образуются при 12–420С, эта способность теряется после длительного выращивания при 42,50С.

1 2 3 4 5 6
Рис. 30. Виды эндопор:
1 – центральная спора, вегетативная клетка не лизируется, 2 – терминальная спора, 3 – «теннисная ракетка», 4 – центральная спора, вегетативная клетка лизируется, 5 – «барабанная палочка», 6 – свободные споры
Споры очень резистентны к физическим (высушиванию, высокой температуре, радиации, вакууму) и химическим (дезинфектантам, токсическим веществам, кислороду) факторам. Это позволяет им выжить в условиях, губительно действующих на вегетативные клетки.
Терморезистентность эндоспор обусловлена:
– практически полным отсутствием свободной воды;
– большим содержанием кальциевой соли дипиколиновой кислоты, которая не встречается у вегетативных клеток;
– особым строением белка;
– особым строением пептидогликана кортекса;
– многослойностью и толщиной оболочки.
Эндоспоры образуются внутри бактериальной клетки. Образование эндоспор у бактерий не является способом размножения. Как правило, внутри бактериальной клетки образуется только одна спора. У отдельных видов рода Clostridium обнаружены клетки с двумя и более спорами.
Объем споры в 10 раз меньше, чем вегетативной клетки. Структурно спора представляет ДНК, окруженную многослойной оболочкой (рис. 31).

Рис. 31. Схематическое изображение строения споры:1 — экзоспориум; 2 — слои споровой оболочки; 3 — внешняя мембрана споры; 4 — кортекс; 5 — внутренняя мембрана споры; 6 — сердцевина
Строение сердцевины спор сходно у разных видов. Объем оболочки споры достигает 50% от всего объема споры. Строение оболочек спор у разных видов сильно различается. Оболочка определяет форму споры, защищает ее от действия литических ферментов и от преждевременного прорастания. Споры мутантных штаммов, лишенные оболочек, прорастают даже в неблагоприятной для роста среде, но проросшие клетки в такой среде погибают.
Споруляция — процесс интенсивного распада белков материнской клетки с одновременным образованием новых белковых компонентов. Поэтому по антигенной структуре споры и вегетативные клетки одного и того же вида различны. Имеются также четкие различия между спорами разных видов. Процесс спорообразования генетически обусловлен. Он длится 18–20 часов, внешние источники питания и энергии не требуются.
Стадии спорообразования (споруляции):
1.Подготовительная. Перед спорообразованием резко снижается синтез ДНК и деление клеток прекращается. Происходит накопление большого количества запасного питательного вещества — гранулезы, откладывающегося в виде гранул. ЦП становится зернистой. Клетки резко увеличиваются в размерах, принимают вид лимона либо барабанной палочки, иногда клетки сохраняют обычную палочковидную форму.
| У спорообразующей бактерии образуется дополнительный нуклеоид, который отходит к одному из полюсов клетки (рис. 32). |
|
Вокруг дополнительного нуклеоида в ЦП спорообразующей клетки образуется спорогенная зона — уплотненный участок ЦП, не имеющий свободной воды.
| 2. Стадия предспоры (проспоры). В ЦПМ, ближе к одному из полюсов клетки, возникает инвагинация. В этом процессе участвуют мезосомы, которые спаивают сближающиеся участки инвагинированных мембран и образуют споровую перегородку (септу), разделяющую клетку на два протопласта, каждый из которых содержит одну хромосому (рис. 33). |
|
Затем материнская клетка поглощает отсеченный участок ЦП с ядром (рис. 34). Это осуществляется путем роста и продвижения периферических участков мембраны материнской клетки по направлению к полюсу клетки. Эта стадия спорообразования обратима.
| Если после образования септы добавить к спорулирующей культуре антибиотик левомицетин, то синтез белков и рост мембраны будет подавлен. Движение периферических участков мембраны, поглощающей отсеченный участок протопласта, будет остановлено. |
|
В результате начавшийся процесс спорообразования превратится в обычный процесс вегетативного деления клеток.
| В конце этой стадии сближающиеся участки мембраны сливаются и меньший из протопластов (проспора) покрывается второй оболочкой, синтезируемой мембраной материнской клетки (рис. 35). |
|
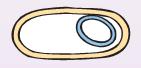
При этом спорогенная зона изолируется от остальной части ЦП. Так внутри клетки возникает новая клетка-проспора, окруженная, в отличие от материнской, двумя трехслойными мембранами — внутренней и внешней. У одних видов проспора остается у полюса клетки, у других видов занимает центральное положение. С этого момента начинается необратимая фаза споруляции.
| 3. Образование кортекса— оболочки между внутренней и внешней мембранами проспоры, состоящей из особых молекул пептидогликана (рис. 36). Кортекс играет защитную роль: он предохраняет сердцевину спор от литических ферментов. |
|
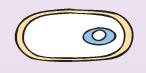
4. Стадия созревания.С внешней стороны наружной мембраны споры образуется внешняя плотная споровая оболочка, в состав которой входят белки, липиды и кальциевая соль дипиколиновой кислоты.
| Снаружи у многих видов анаэробов спора покрывается еще одной дополнительной полипептидной оболочкой — экзоспориумом (рис. 37). |
|
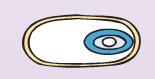
Он имеет вид многослойного чехла, в котором расположена спора. Возникает экзоспориум на ранней стадии формирования споры в виде небольшого пузырька на внешней мембране проспоры, затем пузырек разрастается и охватывает спору со всех сторон. После механического удаления экзоспориума процесс прорастания спор не нарушается.
| 5. Отмирание вегетативной части клетки.На завершающей стадии спорообразования происходит резкое возрастание активности литических ферментов, полностью разрушающих материнскую вегетативную клетку (рис. 38). |
|
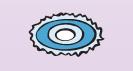
Спора выходит в окружающую среду, где может сохраняться потенциально жизнеспособной тысячи лет (стадия покоя).
Генетический контроль спорообразования. Синтез новых белковых веществ, споровых оболочек, кортекса и экзоспориума осуществляется при активном контроле генома материнской клетки. Роль материнской клетки сводится к тому, чтобы отделить от себя дочернюю клетку, оказавшуюся внутри ее ЦП. Но так как удалить проспору за пределы клеточных границ через ЦПМ и КС стенку невозможно, то материнская клетка изолирует дочернюю от себя, окружая проспору мощными слоями белковой оболочки и переводя ее в анабиотическое состояние. Но это не спасает материнскую клетку от гибели. Хотя она продолжает еще длительное время активно функционировать, даже иногда способна образовать еще одну спору, но все же она не в состоянии вернуться к вегетативному развитию.
Прорастание споры.При попадании в благоприятные условия спора начинает прорастатьв вегетативную клетку. Прорастание длится 4–5 часов, т. е. в 5 раз быстрее, чем спорообразование. Обычно не все споры, вносимые в благоприятную среду, прорастают сразу. Некоторые споры остаются непроросшими в течение различного времени, не теряя своей жизнеспособности.
Способ и время прорастания молодых спор являются постоянными и могут служить видовыми признаками. Если перенести только сформировавшиеся споры в благоприятную питательную среду, их прорастание начинается не сразу, а через 2–3 часа. Старые споры прорастают более длительно.
Стадии прорастания споры:
1. Набухание, связанное с увеличением в споре количества воды.
2. Дерепрессия генома и активация ферментов, участвующих в энергетическом и пластическом метаболизме. На начальной стадии прорастания спор происходит выделение в среду экссудата прорастания. Половину его сухой массы составляет дипиколинат кальция, остальная часть экссудата прорастания представлена свободными аминокислотами, высокомолекулярными и низкомолекулярными белковыми веществами.
Активация ферментов происходит при обработке спор незначительными количествами некоторых веществ (L–аланина, пуриновых нуклеозидов, глюкозы, аденозина), кипячением или пастеризацией, умеренным воздействием ультразвука. Под действием активаторов дипиколиновая кислота, связывающая ферменты, подвергается деполимеризации и неактивная ферментная система превращается в активную.
Напротив, наличие в среде некоторых веществ (мышьяковистых соединений, углекислого натрия, высоких концентраций ряда аминокислот — глицина, метионина, цистеина и валина, D–аланина) угнетает процесс прорастания спор. Механизм угнетающего действия ряда факторов на прорастание спор окончательно неясен.
3. Разрушение плотных оболочек споры под влиянием давления, вызванного ростом. Через образовавшееся отверстие в оболочке споры молодая клетка (проросток, ростовая трубка) выходит в среду и начинает расти (рис. 39). При этом из одной споры образуется одна вегетативная клетка,которая начинает делится (рис. 40).
| 
|
| Рис. 39. Проросток споры | Рис. 40. Юная вегетативная клетка |
Выросты на спорах. Особенностью спорообразования у анаэробов является формирование на спорах выростов разнообразной формы в области контактирующих мембран материнской клетки и проспоры. Каждому виду анаэробных бактерий свойствен свой тип строения выростов, этот признак наследственно закреплен и очень устойчив. Выросты состоят в основном из белка, в веществе выростов также обнаружены полисахариды и липиды.
Предположения о функции выростов на спорах:
– ответственны за контакт клеток в системе спорангий — спора,
– подают споре при благоприятных условиях «команду» на прорастание,
– играют важную роль в созревании спор, участвуют в формировании споровых покровов и кортекса,
– являются результатом нарушений в нормальном обмене веществ.
Колпачки на спорах. На спорах анаэробов образуются колпачки ячеистого строения. Ячейки — мешочки, заполненные газообразным веществом. Функция колпачков — увеличение подвижности и плавучести спор в водной среде, что способствовует их распространению в природе. Подобно понтонам ячеистые колпачки не позволяют спорам тонуть. Споры с колпачками не удается осадить даже при центрифугировании.
Функции эндоспор:
1. Способность бактерий образовывать споры позволяет сохранить наследственную информациюпри попадании вегетативной клетки в неблагоприятные условия внешней среды и способствует сохранению вида. Споры могут оставаться жизнеспособными в течение десятков, сотен и даже тысяч лет. Жизнеспособные споры были выделены из египетских мумий, трупов мамонтов.
2. Длительное существование спор бактерий в покоящемся состоянии в окружающей среде имеет существенное эпидемиологическое значение. Попадание спор патогенных бактерий в глубокие колотые и резанные раны, в которых создается анаэробиоз, может привести к возникновению анаэробной раневой инфекции (столбняка или газовой гангрены).
3. Способность бактерий образовывать споры, различающиеся по форме, размерам и локализации в клетке, является таксономическим видовым признаком, который используется для идентификации.
Форма спор может быть овальной, шаровидной, яйцевидной, цилиндрической.
Размер спор у бацилл приблизительно равен диаметру клетки, поэтому клетка не меняет формы. У клостридий размер спор превышает диаметр клетки, поэтому клетка расширяется в середине и принимает форму веретена.
Расположение спор в клетке может быть центральное (у сибиреязвенной бациллы), терминальное (у клостридий столбняка), субтерминальное (у клостридий ботулизма и газовой гангрены).
Выявление эндоспор:
1. При обычных способах окраски споры не прокрашиваются и имеют вид неокрашенных пустот внутри окрасившихся вегетативных клеток, так как плотная оболочка спор непроницаема для воды. Споры видны благодаря своему высокому показателю преломления — такому же как у обезвоженного белка. Это указывает на содержание в спорах бактерий большого количества богатого белком материала, сконцентрированного в малом объеме. Спора содержит почти все сухое вещество материнской клетки, но занимает в 10 раз меньший объем.
2. Специальные методы окраски используют в сомнительных случаях. При этом используют протравы, которые разрыхляют оболочку споры и облегчают проникновение красителя. Окрасившиеся споры обладают кислотоустойчивостью в отличие от вегетативного тела микробной клетки, обесцечивающегося под действием кислоты.
Окраска по Ожешко:
| – на высушенный нефиксированный мазок (толстый, на краю стекла) наносят несколько капель 0,5 % HCl и подогревают 1–2 мин до закипания, остатки кислоты сливают; – остывший препарат промывают водой, подсушивают, фиксируют на пламени горелки; далее красят как по Цилю-Нильсену, для докрашивания можно использовать 1 % малахитовый зеленый. Окрасившиеся споры (рубиново-красные) обладают кислотоустойчивостью в отличие от вегетативных тел микробных клеток (синих или зеленых) (рис.41). |
|
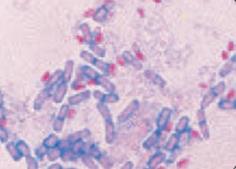
3. Фазово-контрастная микроскопия.
4. Электронная микроскопия.
Экзоспоры, в отличие от эндоспор, образуются вне бактериальной клетки и являются способом размножения у актиномицетов. На одну бактериальную клетку приходится не одна, а много спор. Экзоспоры менее устойчивы во внешней среде.
Дата добавления: 2021-02-19; просмотров: 1180;
Поиск по сайту
Узнать еще
- ATP-зависимые протеазы прокариот
- VIII. 2. Большой круговорот вещества и роль в нем человека
- Абиотическая миграция вещества
- Аварийно химически опасные вещества АХОВ)
- Аварийно-химические отравляющие вещества
- Агрегатные состояния вещества
- Агрегатные состояния вещества.
- Азотосодержащие экстрактивные вещества

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
