Участвуют в спорообразовании.
Выявление мезосом:
1. Электронная микроскопия.
ЦИТОПЛАЗМА (ЦП)
Строение.Цитоплазма (протоплазма)—содержимое клетки, окруженное ЦПМ и занимающее основной объем бактериальной клетки. ЦП является внутренней средой клетки и представляет собой сложную коллоидную систему, состоящую из воды (около 75%) и различных органических соединений (белков, РНК и ДНК, липидов, углеводов, минеральных веществ).
Располагающийся под ЦПМ слой протоплазмы более плотный, чем остальная масса в центре клетки. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: нуклеоидом, плазмидами, рибосомами и включениями.
Функции цитоплазмы:
1. Содержит клеточные органеллы.
Выявление цитоплазмы:
1. Электронная микроскопия.
НУКЛЕОИД
Строение. Нуклеоид— эквивалент ядра эукариот, хотя отличается от него по своей структуре и химическому составу. Нуклеоид не отделен от ЦП ядерной мембраной, не имеет ядрышек и гистонов, содержит одну хромосому, имеет гаплоидный (одиночный) набор генов, не способен к митотическому делению.
Нуклеоид расположен в центре бактериальной клетки, содержит двунитевую молекулу ДНК, небольшое количество РНК и белков. У большинства бактерий двунитевая молекула ДНК диаметром около 2 нм, длиной около 1 м с молекулярной массой 1–3х109 Да замкнута в кольцо и плотно уложена наподобие клубка. У микоплазм молекулярная масса ДНК наименьшая для клеточных организмов (0,4–0,8x109 Да).
ДНК прокариот построена так же, как и у эукариот (рис. 25).

Рис. 25. Строение ДНК прокариот:
А — фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание: 1 — цитозин; 2 — гуанин.
Б — двойная спираль ДНК: Д — дезоксирибоза; Ф — фосфат; А — аденин; Т — тимин; Г — гуанин; Ц — цитозин
Молекула ДНК несет множество отрицательных зарядов, так как каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками — гистонами. В клетках прокариот гистонов нет, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами и ионами Mg2+.
По аналогии с хромосомами эукариот бактериальная ДНК часто обозначается как хромосома. Она представлена в клетке в единственном числе, поскольку бактерии являются гаплоидными. Однако перед делением клетки число нуклеоидов удваивается, а во время деления увеличивается до 4 и более. Поэтому термины «нуклеоид» и «хромосома» не всегда совпадают. При действии на клетки определенных факторов (температуры, pH среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках обнаруживается по одной копии хромосомы.
Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Специальные исследования показали, что хромосомы прокариот — высокоупорядоченная структура. Часть ДНК в этой структуре представлена системой из 20–100 независимо суперспирализованных петель. Суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По периферии нуклеоида располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК). Поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами.
Кроме нуклеоида в цитоплазме бактериальной клетки могут находиться плазмиды — факторы внехромосомной наследственности в виде дополнительных автономных кольцевых молекул двунитевой ДНК с меньшей молекулярной массой. В плазмидах также закодирована наследственная информация, однако она не является жизненно необходимой для бактериальной клетки.
Функции нуклеиода:
1. Хранение и передача наследственной информации, в том числе о синтезе факторов патогенности.
Выявление нуклеоида:
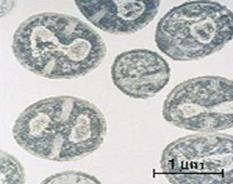
|
| Рис. 26. Нуклеоид стафилококка (трансмиссивная электронная микроскопия) |
1. Электронная микроскопия: на электронограммах ультратонких срезов нуклеоид имеет вид светлых зон меньшей оптической плотности с фибриллярными, нитевидными структурами ДНК (рис. 26). Несмотря на отсутствие ядерной мембраны, нуклеоид довольно четко отграничен от цитоплазмы.
2. Фазово-контрастная микроскопия нативных препаратов.
3. Световая микроскопия после окраски специфическими для ДНК методами по Фельгену, по Пашкову или по Романовскому-Гимза:
– препарат фиксируют метиловым спиртом;
– на фиксированный препарат наливают краситель Романовского-Гимза (смесь равных частей трех красок — азура, эозина и метиленового синего, растворенных в метаноле) на 24 часа;
– краску сливают, промывают препарат дистиллированной водой, высушивают и микроскопируют: нуклеоид окрашивается в фиолетовый цвет и располагается диффузно в цитоплазме, окрашенной в бледно-розовый цвет.
РИБОСОМЫ
Строение.В отличие от клеток эукариотов рибосомы бактерий не объединены в эндоплазматическую сеть и имеют меньшую молекулярную массу.
Бактериальные рибосомы — рибонуклеопротеиновые частицы размером 20 нм, состоящие из двух субъединиц 30S и 50S (рис. 27).
| Малая субъединица 30S содержит 16S рРНК и 21 белок; большая субъединица 50S — 23S и 5S рРНК и 34 белка. Перед началом синтеза белка происходит объединение этих субъединиц в одну — 70S (S — константа седиментации, единица Сведберга, характеризующая скорость осаждения в центрифуге при стандартных условиях). | 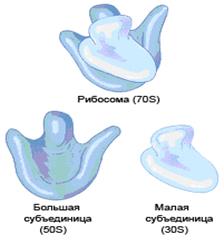
|
| Рис. 27. Бактериальная рибосома |
Общая масса рибосом составляет примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) — 80–85% всей бактериальной РНК.
Количество рибосом в бактериальной клетке зависит от интенсивности белкового синтеза и колеблется от 5000 до 90 000. Число рибосом тем больше, чем быстрее растет клетка.
В процессе белкового синтеза в клетке образуются полирибосомы (полисомы) — структуры, состоящие из большого числа рибосом, молекул информационной и транспортных РНК. Полисомы могут находиться в цитоплазме или быть связанными с мембранами клетки. По окончании синтеза белка полисомы вновь распадаются на одиночные рибосомы.
Функции рибосом:
1. Синтез белка. Существует два основных типа рибосом — 70S (у прокариот) и 80S (у эукариот). Бактериальные рибосомы, отличные от белоксинтезирующих систем эукариотических клеток, являются «мишенью» для действия многих антибиотиков. Некоторые антибиотики полностью или частично подавляют синтез белка, протекающий на рибосомах 70S, но не затрагивают функцию рибосом 80S. Так как наружная мембрана митохондрий (митохондрии содержат рибосомы 70S) очень мало проницаема, в терапевтических концентрациях антибиотики не действуют на эукариотические клетки. Деление органелл у эукариот прекращается лишь при использовании в 1000 раз более высоких концентраций антибиотиков.
Выявление рибосом:
1. Электронная микроскопия.
ВКЛЮЧЕНИЯ
Строение.У многих микроорганизмов при избытке питательных веществ в окружающей среде в ЦП клеток откладываются включения – запасные вещества разной химической природы и функционального назначения.
Включения представлены полисахаридами, липидами, полифосфатами, полипептидами, отложениями минеральных веществ (серы, кальция, железа)
(табл. 6).
Изполисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобное вещество — гранулеза — специфический запасной полисахарид анаэробных споровых бактерий группы клостридий. Названные полисахариды построены из остатков глюкозы. В неблагоприятных условиях они используются в качестве источника углерода и энергии.
Липиды накапливаются в виде гранул, резко преломляющих свет и поэтому хорошо различимых в световой микроскоп. Запасным веществом такого рода является полимер β–оксимасляной кислоты, накапливающийся в клетках многих прокариот. У некоторых бактерий, окисляющих углеводороды, поли–β–оксимасляная кислота составляет до 70% сухого вещества клеток. Отложение липидов в клетке происходит в условиях, когда среда богата источником углерода и бедна азотом. Липиды служат для клетки хорошим источником углерода и энергии.
Полифосфаты, содержащиеся в волютиновых гранулах, используются клетками как источник фосфора. Полифосфаты содержат макроэргические связи и являются депо энергии, хотя их роль как источника энергии незначительна.
Все запасные вещества представлены в виде высокомолекулярных полимерных молекул, в ряде случаев отграниченных от ЦП белковой мембраной, т. е. находятся в осмотически неактивном состоянии и нерастворимы в воде.
Включения — продукты клеточного метаболизма, имеющие приспособительное значение. Когда в этих веществах возникают потребности, они снова включаются в метаболизм.
Таблица 6
Дата добавления: 2021-02-19; просмотров: 1084;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
