Анатомические основания рефлекторных путей
Согласно вышеприведенному заключению, простейший и кратчайший рефлекторный путь или так называемая рефлекторная дуга у позвоночных животных должна состоять из трех нейронов:
1) афферентных, или чувствительных, нейронов, которые приносят периферические импульсы от рецепторов в центральную нервную систему,
2) промежуточных, которые проводят эти импульсы к двигательным нейронам, и
3) эфферентных, или двигательных, нейронов, которые передают их далее периферическому двигательному органу, так называемому эффектору.
На рис. 22 дана схема рефлекторной дуги координированных рефлексов сгибания, разгибания и рефлекса потирания с точки зрения нейронной теории. По этой теории вся рефлекторная реакция с возбуждением одних мышц и торможением других осуществляется путем проведения в спинном мозгу афферентных импульсов возбуждения через нервные клетки и их аксоны.
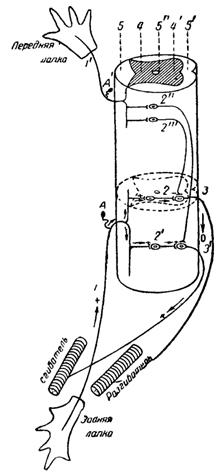
Рис. 22. Схематическое изображение рефлекторных дуг у лягушки согласно нейронной теории. Изображены: часть спинного мозга, кожа на лапке, мышцы-антагонисты и нервные связи между ними. 1 — чувствительный нейрон от воспринимающего поля задней конечности; 2, 2' — промежуточные нейроны спинного мозга, которые производят координированную иннервацию в рефлексе сгибания; 3 — двигательный нейрон разгибателя; 3' — двигательный нейрон сгибателя.
Плюсы на этих нейронах обозначают возбуждение, минусы — торможение; стрелка везде — направление процесса. 1’ — чувствительный нейрон воспринимающего поля потирания передних конечностей; 2', 2" — координирующие нейроны рефлекса потирания: возбуждение и торможение этой дуги не указаны. А, А' — клетки чувствительных нейронов; 4 — задний рог серого вещества спинного мозга; 4'—передний рог этого же вещества; 5 — задние столбы винного мозга; 6' — передние столбы
На рис. 23 дана схема рефлекторной дуги тех же рефлексов, согласно нейронно-нейропильной теории. Здесь коллатерали заднекорешковых волокон, затем клетки и аксоны промежуточных нейронов служат для проведения афферентных импульсов возбуждения к мышцам, а нейропиль — для торможения их.
На непременное участие промежуточных нейронов в рефлекторных реакциях указывают как старые, так и новейшие гистологические исследования. Так, например, после перерезки задних корешков кошки на клетках промежуточных нейронов оказывается гораздо больше дегенерированных окончаний заднекорешковых волокон, чем на клетках двигательных (Гофф, 21).
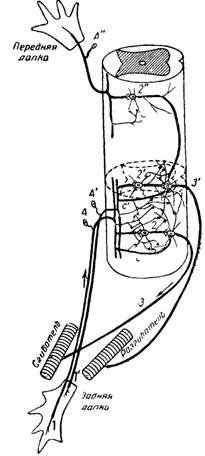
Рис. 23. Схематическое изображение рефлекторных путей по нейронно-нейропильной теории. Те же афферентные нейроны и двигательные нейроны, что и на рис. 22, связываются между собой не только через промежуточные нейроны, но и через нейропиль, образуемый дендритами и коллатералями нейритов всех этих клеток. От задней лапки идут два афферентных нейрона: один—для сгибательного рефлекса (ІА), а другой — для разгибательного (1' А').
Когда возбуждается первый афферентный нейрон, то через промежуточный нейрон (2), а также через коллатераль с возбуждается сгибатель, а разгибатель через нейропиль тормозится. Когда же возбуждается второй афферентный нейрон, то через промежуточный нейрон (2’) н коллатераль с' возбуждается разгибатель, а сгибатель через нейропиль тормозится. От передней лапки идет один афферентный нейрон (А"): он через вставочный (2*) нейрон возбуждает разгибатель, а через нейропиль тормозит сгибатель
Кроме того, известно, что каждый чувствительный нейрон, проходя по задним столбам спинного мозга, отдает свои коллатерали не в одном сегменте, а в целом ряде сегментов как впереди места вхождения, так и позади него. Вследствие этого каждое рецептивное поле и каждый чувствительный нерв имеют возможность влиять почти на всю центральную нервную систему.
Правда, каждый чувствительный нейрон связан особенно интимно с двигательными нейронами соответствующего сегмента и легче всего влияет на эти нейроны. Но факты показывают, что при определенных условиях данные чувствительные нейроны способны влиять и на двигательные нейроны других сегментов. Кроме того, промежуточные нейроны связаны не с одним каким-либо комплексом двигательных нейронов.
Они связаны также более или менее интимно с двигательными нейронами разных отделов мозга. Известно, например, что дыхательный центр, который лежит в продолговатом мозгу, при определенных условиях в состоянии возбудить всю мускулатуру тела. Следовательно, не может быть речи о строго изолированных нервных путях для рефлекторных реакций.
Каково бы ни было раздражение, и где бы оно ни происходило, вызываемое им возбуждение должно выходить из пределов соответствующей рефлекторной дуги и захватывать в большей или меньшей степени остальную нервную систему.
Обычно представляют себе, что передача возбуждения с афферентного нервного волокна на промежуточный и двигательный нейроны происходит через клетки этих нейронов. Но такое представление отнюдь не обязательно. Афферентный и другие нейроны оканчиваются не только на теле клетки, но н на том холмике, где начинается аксон клетки. Значит, возбуждение может передаться с нейрона на нейрон и без участия самого тела клетки. А опытами на ганглии мантии одного из головоногих моллюсков (Loligo vulgaris) было доказано, что на самом деле синаптическая передача возбуждения может происходить без участия клеточных тел.
Здесь из гигантских клеток, расположенных на краю ганглия, выходят гигантские нервные волокна к мышечным волокнам. От этих волокон еще в ганглии отходят коротенькие коллатерали, которые вступают в синаптические контакты с окончаниями нервного волокна, идущего из центральной нервной системы. Если отрезать клеточную часть, то ганглий вое же продолжает проводить возбуждение от пресинаптического волокна на постсинаптическое.
Это указывает, что ядро, тельца Ниссля, аппарат Гольджи и другие составные части клетки не являются необходимыми для осуществления передачи возбуждения с нейрона на нейрон (Юнг, 22).
Если нельзя говорить о строго локализированных рефлекторных дугах, то тем более нельзя признавать существование строго обособленных координирующих аппаратов в спинном мозгу. При некоторой нормальной возбудимости центральной нервной системы каждый чувствительный нерв вызывает координированную реакцию через тот отдел мозга, куда входит данный нерв непосредственно.
Но при некоторой повышенной возбудимости в данных координирующих аппаратах последние будут вовлекаться в работу и при раздражении других чувствительных нервов. Более того, при нормальной возбудимости центральной нервной системы данный чувствительный нерв вызывает реакцию исключительно через определенный комплекс промежуточных, координирующих нейронов; при некоторой повышенной возбудимости двигательных клеток данный нерв вызывает реакцию и помимо координирующего аппарата, прямо из двигательных клеток, и притом тем сильнее, чем они анатомически ближе к месту вхождения чувствительного нерва (Беритов, 9).
Отсюда следует, что фактически нет строго обособленного координирующего центра, который действовал бы исключительно под влиянием определенных периферических раздражений. Характер внешней реакции на данное раздражение определяется главным образом функциональным состоянием всей центральной нервной системы в момент раздражения, а не просто анатомическими соотношениями между рецептором и центральной нервной системой.
Дата добавления: 2023-04-03; просмотров: 1367;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
