Некоторые функциональные отделы коры. Первичные моторные и сенсорные передаточные поля
Каждая функциональная часть мозговой коры человека может рассматриваться как происшедшая из (или являющаяся наружной проекцией) определенных полей серого вещества старого мозгового ствола (см. рис. 4).
Таким образом, проекционное поле во вновь сформированной коре, по-видимому, служит усилению и расширению функции, которая также выполняется каким-то рудиментарным образом старым мозгом более элементарных животных.
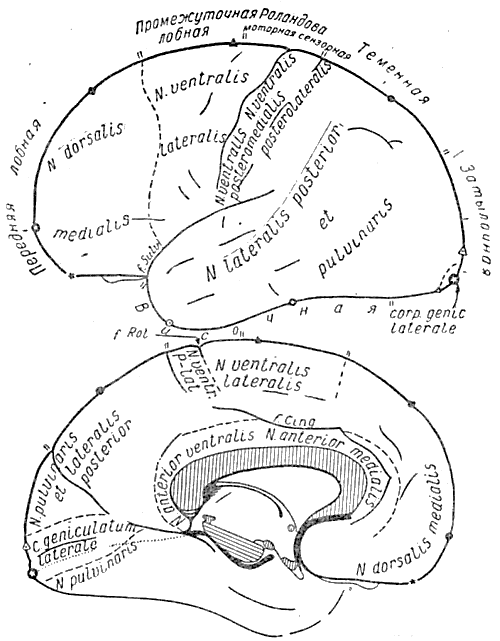
Рис. 4. Проекция ядер таламуса и коленчатых тел на кору больших полушарий, составленная на основе учета таламокортикальных связей по данным работы Эрла Уолкера. (1938а, 19386). Латеральная поверхность полушарий показана на верхнем рисунке, медиальная — на нижнем
Например, передние отделы лобной коры могут рассматриваться как результат развития дорсо-медиального ядра зрительного бугра, а большая часть височной коры как наружная проекция подушки и задней части латерального ядра зрительного бугра (см. рис. 3). Это во многих отношениях более верный указатель функционального подразделения коры по сравнению с цитоархитектоническим расчленением ее (например, полями Бродмана, рис. 5).
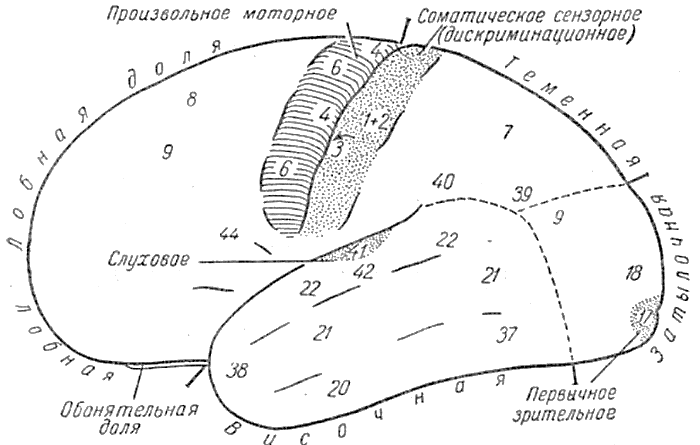
Рис. 5. Левое полушарие с некоторыми архитектоническими корковыми подразделениями по Бродману (показаны цифрами). На рисунке специально отмечены (пунктиром, либо сплошными линиями) такие участки, как например, некоторые корковые передающие станции движений и чувствительности. Вкусовая и висцеральная чувствительность, станции которых расположены глубоко в сильвиевой борозде и островке, не отмечены на рисунке, так же как и вестибулярная чувствительность, которая, возможно, связана с небольшим участком коры вблизи слухового поля.
Большая часть слухового поля спрятана в сильвиевой борозде; большая часть первичного зрительного поля в шпорной борозде, соматического поля дискриминативной чувствительности — в роландовой борозде. Пути, которые обслуживают болевую чувствительность, не показаны: они оканчиваются в таламусе н не имеют захода к коре, имеющейся в путях других видов чувствительности
Полезным вступлением к нашему обсуждению речи явится описание в общих чертах некоторых функциональных полей мозговой коры человека. Эти поля определялись, главным образом, с помощью электрического раздражения или оперативной экстирпации. Поля, которые выявлялись таким образом, являются сенсорными, моторными или психическими.
Первичные моторные и сенсорные передаточные поля. Первичное моторное передаточное поле мозговой коры расположено в передней центральной извилине, распространяясь в значительной степени в сторону центральной роландовой борозды. Здесь берет свое начало часть кортикоспинального тракта. Но поток нервных импульсов, обуславливающих произвольную деятельность, возникает не в коре, а субкортикально (Пенфильд, 1954а).
Что это так, подтверждается тем фактом, что удаление серого вещества непосредственно впереди или кзади от передней центральной извилины не лишает больного возможности контролировать движения противоположной руки, и управлять ею соответственно многочисленным источникам информации (рис. 6).
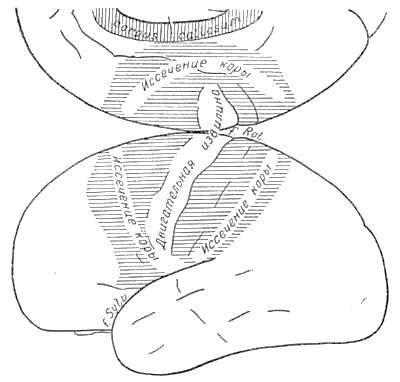
Рис. 6. Схема показывает поля (сплошные линии), прилегающие к передней центральной извилине, удаление которых у человека не лишает его способности осуществлять произвольные движения с помощью различных источников сенсорной информации, поступающей в мозг через оба полушария
Он может, по-прежнему, направлять движения этой руки в соответствии со зрительной информацией, которая поступает в мозг через зрительное поле в затылочной доле той же стороны. Если затылочная доля на этой стороне удалена, больной конечно может направлять руку, благодаря зрительным импульсам, которые поступают в мозг через другую затылочную долю.
Поэтому кажется более обоснованным предположение, что поток импульсов (пунктирная линия на рис. 7), определяющий движение руки, может происходить из ганглионарных полей центрэнцефалической системы. Поскольку руки так часто действуют одновременно, можно полагать, что ганглионарное поле имеет интимное отношение к формированию потоков импульсов, двигающихся в разных направлениях к моторным полям, связанным с контролем руки в противоположном полушарии.
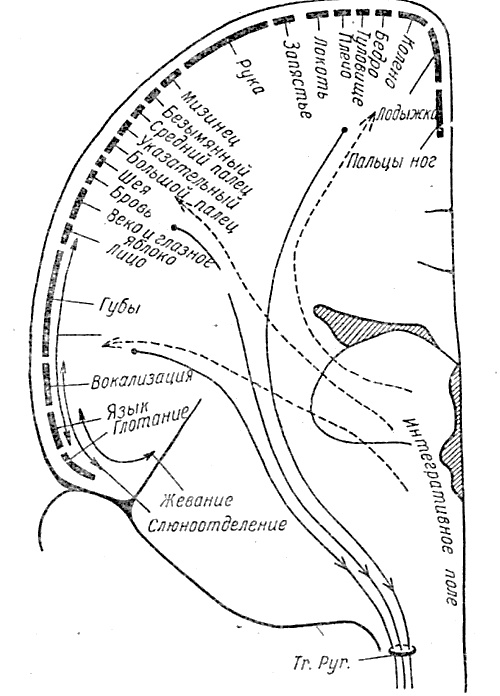
Рис. 7. Пути произвольных движений. Поперечное сечение через правое полушарие вдоль передней центральной извилины. Пунктирными линиями показаны проводящие пути управления произвольными движениями, начинающиеся в сером веществе где-то в верхнем мозговом стволе и идущие к двигательным передаточным областям в передней центральной извилине.
Далее, как это показано сплошными линиями, они идут вниз по кортико-спинальному тракту в направлении к мышцам. Последовательность ответов на электрическое раздражение поверхности коры (сверху вниз, вдоль двигательной области от пальцев ног, затем руки и лица до глотки) не изменяется у разных индивидуумов. (По Пенфильду и Джасперу, 1954)
Моторное иоле коры является, следовательно, «платформой прибытия и отправления импульсов» (см. рис. 7). Его функция заключается в проведении и возможном преобразовании с помощью вторичных моторных полей потока импульсов определенного характера, которые возникают в центрэнцефалической системе и предназначены для скелетных мышц. Имеются поэтому супра- или прекортикальные части проводящих путей для импульсов произвольных движений и инфра- или посткортикальные части, которые объединяются в той области коры, которая связана с передачей моторных импульсов.
Соматическое сенсорное поле задней центральной извилины также является передаточной областью. Удаление коры сзади от него или даже в передней центральной извилине — впереди от него не лишает больного информации, которая нормально доставляется на эту «платформу прибытия». Поток соматических сенсорных импульсов, который приходит из кожи, мышц и суставов, направляется в заднюю центральную извилину (рис. 8) после ганглиозного перерыва в латеральном ядре зрительного бугра. Для соединения с центрэнцефалической системой этот поток должен возвратиться назад по посткортикальному пути (пунктирная линия на рис. 8).
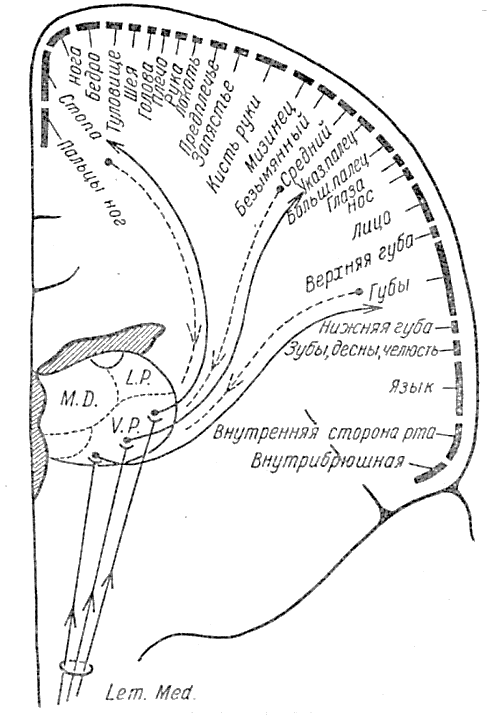
Рис. 8. Соматическая чувствительность. Поперечное сечение через левое полушарие вдоль задней центральной извилины. Афферентные проводящие пути для дискриминативной чувствительности показаны сплошной линией, поднимающейся через медиальную петлю к передаточным областям задней центральной извилины Затем, как это показано пунктирной линией, они входят в центрэнцефалические цепи интеграции.
Между сенсорной и двигательной активностью имеются, без сомнения. взаимосвязи, элементы которых в области центральной борозды показаны на этом н предшествовавших рисунках. (По Пенфильду и Джасперу, 1954)
Примерно то же можно сказать о первичном зрительном поле коры. Оно локализуется по сторонам шпорной борозды, расположенной на медиальной поверхности затылочной доли (см. рис. 2—9). Удаление этого поля вызывает полную слепоту в противоположном поле зрения (гомонимная гемианопсия) за исключением, может быть, небольшого полулунного диска центрального зрения. Из этого участка в центре поля зрения афферентный поток импульсов идет (как считают некоторые исследователи) через зрительное поле как ипсилатеральной коры, так и контралатеральной.
Удаление прилежащей части затылочной области коры, если первичная шпорная область сохранена, не вызывает у больного слепоты в контралатеральном поле зрения, а также затруднений в зрительном контроле движений конечностей.
Дата добавления: 2023-03-24; просмотров: 1356;
Поиск по сайту
Узнать еще
- Вторичные и дополнительные поля
- Головной мозг. Проекционные поля коры
- Каковы функциональные особенности нейрона? Нейроглия и ее функции
- Некоторые проблемы вокальной педагогики
- Некоторые свойства ионизирующих излучений. Радиоактивность и ядерные реакции
- Отделы, мозговой коры, выявляемые с помощью раздражения
- Что называется нейроном, их классификация. Каковы основные структурно-функциональные элементы нейрона?

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
