Общее свойство рецепторов
Общее свойство рецепторов. Как указывалось выше, одни рецепторы представляют собой свободные окончания чувствительных нервных волокон; другие имеют более или менее сложное строение с участием специальных клеток, на которых оканчиваются чувствительные нервные волокна. В тех случаях, когда рецепторы являются свободными окончаниями чувствительного волокна, адекватное раздражение действует непосредственно на эти окончания.
Таковы свободные окончания чувствительных волокон в эпителии эпидермиса, в сухожилиях, в склере, в соединительно-тканных прослойках мышечных оболочек всех внутренних органов. Свободные окончания образуют своеобразные нейрофибриллярные расширения, петельки, пластинки. В основном они являются механорецепторами, возбуждаясь при всяких растяжениях и сокращениях органов, а в определенных случаях, видимо, они же дают импульсы, воспринимающиеся как боль (Лаврентьев, 19).
Когда рецептор имеет сложное строение с участием специальных клеток, адекватное раздражение действует на эти специальные клетки. Таковы все рецепторы давления и прикосновения, все хеморецепторы,— например органы вкуса, множество рецепторов кровеносных сосудов, кишечника, почек и других внутренних органов. Процессы, возникающие в этих специальных клетках под (влиянием раздражения, являются источником раздражения для окончаний чувствительного волокна (Лаврентьев, 19).
Физиологическая деятельность каждого рода рецепторов более или менее одинакова. Каждый рецептор в ответ на адекватное — т. е. со ответствующее ему — раздражение отвечает рядом (возбуждений. Это наблюдается в сетчатке глаза при раздражении светом, в кожных рецепторах — при давлении, при сотрясении одного волоска, в мышечных рецепторах — при растяжении мышц. Возникающие здесь импульсы возбуждения по чувствительным нервным волокнам передаются центральной нервной системе.
Чем сильнее раздражение, тем выше ритм возбуждения рецепторов и тем дольше он держится. Интенсивность импульсов возбуждения, получаемых от одного рецептора, одна и та же (Эдриан, 6). Следовательно, деятельность рецепторов протекает согласно закону возбуждения; каждый раз рецептор возбуждается с участием всей возбудимой системы. Но, в отличие от нервного волокна, возбуждение рецептора после однократного раздражения повторяется много раз друг за другом.
Очевидно, после одного раздражения в рецепторах создаются все необходимые физико-химические условия для того, чтобы после первого изохронного расщепления и последующего восстановления возбудимой системы вновь многократно происходило изохронное расщепление и восстановление этой системы без каких- либо добавочных внешних раздражений.
Продолжительность отдельных импульсов в чувствительных волокнах от кожных рецепторов различна, В случае возбуждения тактильных рецепторов лягушки она варьирует между 0.5 и 1.2 σ, при болевых раздражениях она значительно больше; в некоторых случаях болевые импульсы продолжаются до 10 σ (Дзидзишвили, 20).
Многие рецепторы разряжаются импульсами возбуждения при полном отсутствии внешних раздражений, т. е. спонтанно. Таковы сетчатка, кортиев орган, статический орган. Зрительный нерв, например, обнаруживает импульсы возбуждения при абсолютной темноте, а VIII нерв — при абсолютной тишине, что доказывается путем отведения биотоков от соответствующих чувствительных нервов.
Это происходит, по-видимому, потому, что физико-химические условия в межуточной (межклеточной) среде и внутри рецептивных клеток не вполне одинаковы, вследствие чего межуточная среда является постоянным источником раздражений для чрезвычайно чувствительных рецептивных клеток (Беритов и Цкипуридзе, 17).
Высокой чувствительностью рецептивных клеток, собственно говоря, обусловливается то явление, что в ответ на каждое кратковременное раздражение они. разряжаются длительным рядом нервных импульсов. Итак, многим рецепторам свойственна спонтанная ритмическая деятельность.
Интенсивность и ритм нервных импульсов в чувствительных волокнах при раздражении разных рецепторов довольно разнообразны. Особенно редки и слабы импульсы болевых рецепторов (рис. 18). Высший ритм возбуждения болевых рецепторов очень низок. По данным Эдриана, при самых сильных болевых раздражениях (раздавливание и прижигание кожи) высший ритм возбуждения этих волокон не превышает 20 в 1 сек (7). С этим свойством рецепторов хорошо согласуется скорость проведения и вообще физиологические особенности отходящих от них чувствительных волокон.
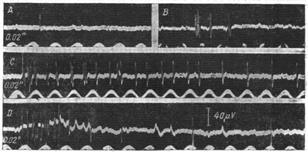
Рис. 18. Афферентная импульсация при кожных раздражениях. В шлейфный осциллограф отводятся импульсы возбуждения дорзальных нервов лягушки, возникающие при точечном раздражении кожи едины. Отведение в он. А монополярное, в остальных опытах биполярное. А, В и С — эффект тактильного раздражения различных точек.
Несмотря на то, что интенсивность раздражения во всех этих случаях одинакова (прикосновение кончиком волоса), адаптация наступает в различное время: в случае А появился один импульс, в случае В — кратковременная группа импульсов, а на С — показано начало импульсации, которая длилась 0.8". D — эффект укола иглой. Показано начало записи, где наряду с быстрыми импульсами видны медленные болевые импульсы. Время 0.02 сек. (Дзидзишвшш)
Как уже указывалось, скорость проведения возбуждения в чувствительных волокнах болевых рецепторов много меньше, чем в чувствительных волокнах тактильных рецепторов. В начале раздражения ритм возбуждения этих болевых рецепторов медленно нарастает до максимума; кроме того, импульсы возбуждения продолжают возникать длительное время и по прекращении раздражения (Эдриан, 10). Это — характерное отличие болевых рецепторов от рецепторов других видов.
Импульсы чувствительных волокон, идущих от рецепторов прикосновения, особенно сильны и часты, причем они наиболее часты в самом начале раздражения этих рецепторов и прекращаются сейчас же по окончаний раздражения. Так, например, если направить на тактильные рецепторы струю воздуха, прерываемую зубчатым колесом 250—300 раз, то в отходящих от них нервных волокнах можно наблюдать 250—300 импульсов в 1 сек., т. е. по одному импульсу возбуждения от каждого дуновения (Каттел и Хоглэнд, 16).
Реакции мышечных рецепторов характеризуются еще более высоким ритмом. При некоторых малых растяжениях высший ритм возбуждения чувствительных нервов невысок, но при больших растяжениях он сильно нарастает, доходя до 290 в 1 сек. у лягушек н до 500 в 1 сек. у теплокровных (Мэтьюс, 8), и даже до 1000 в 1 сек. (Ллойд, 18). Толщина мышечных чувствительных волокон также характерно отличается от кожных; толщина первых 12—20 μ, а вторых — 6—12 μ (Ллойд, 18).
Рис. 18 и 19 иллюстрируют ритмическое течение возбуждения рецепторов, На рис. 18 представлены электрограммы чувствительного нерва при раздражениях кожных рецепторов лягушки. На рис. 19 дана картина токов возбуждения чувствительных нервов при раздражении одного мышечного рецептора путем растяжения.

Рис. 19. Биоэлектрические токи возбуждения чувствительного мышечного нерва при растяжении мышцы на пальце лягушки. Записывается осциллографом Мэтьюса. Температура 17е С. А — в момент растяжения тяжестью в 5 г; Б — 2 сек. позднее; С — 5 сек. позднее; D — та же реакция, записанная при большой скорости фотографического аппарата. На первых трех рисунках время по 0.2 сек., а на В—0.01 сек. Токи возникают с большой регулярностью и почти одинаковой высоты (Мэттьюс)
Дата добавления: 2023-04-03; просмотров: 1340;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
