Анатомическая локализация координирующих аппаратов
Роль задних и передних корешков в рефлекторной реакции спинного мозга. Все чувствительные волокна от рецепторов конечностей, шеи и туловища вступают в спинной мозг через задние корешки спинного мозга. Чувствительные волокна одного корешка начинаются в определенной кожно-мышечной области, как это, например, показано для поясничных задних корешков лягушки на рис. 17.
Передние корешки проводят возбуждение из спинного мозга на периферию. Они производят сокращение мышц и потому называются двигательными. Несмотря на то, что передние корешки своими многочисленными дендритами стоят в связи с разными центральными элементами мозга, а их волокна имеют коллатерали, которые также заканчиваются в сером веществе мозга, раздражением одного двигательного корешка нельзя вызвать внешних двигательных реакций через другие двигательные корешки, т. е. рефлекторных реакций.
Последние можно вызвать только раздражением задних корешков или задних и боковых столбов, где проходят чувствительные волокна. На этом основании был сформулирован закон одностороннего проведения возбуждения в центральной нервной системе от чувствительных нейронов к двигательным.
Значение спинномозговых узлов в координации рефлекса. Прежде чем возбуждение дойдет от рецептивного поля до спинного мозга, оно должно пройти через спинномозговой нервный узел, где находятся клетки чувствительных нейронов. Из каждой такой клетки выходит один нейрит, который в области узла делится на две ветви. Одна ветвь идет к рецептивному полю и там оканчивается в рецепторе; другая вступает в спинной мозг, где дает много боковых ветвей, так называемых коллатералей. Эти коллатерали оканчиваются здесь же или в головном мозгу. Следует решить, какую роль играет этот узел в рефлекторной деятельности.
Опыт Штейнаха с удалением чувствительных клеток из спинномозгового узла. В спинномозговых узлах лягушки клетки расположены группой в одном месте, именно на дорзальной стороне. Штейнаху удавалось удалить эту часть так, что при этом не повреждались другие части узла. Эта операция не прекращала рефлекторного действия через данный узел. В продолжение нескольких дней рефлекс вызывался раздражением того воспринимающего поля, чувствительные нервы которого проходили через оперированный узел.
Гистологическая картина после операции была такова: часть клеток вовсе не существовала, остальная часть была перерождена, дегенерирована; нейриты же этих клеток более или менее уцелели. Отсюда было сделано заключение, что импульсы возбуждения от воспринимающего поля могут проходить в узле без участия клеток, что они могут с периферической ветви нейрита перейти непосредственно на центральную.
Если раздражать чувствительный нерв, а его продолжение до и после узла поочередно отводить в цепь гальванометра или осциллографа, то ритм, а также и общий ход биоэлектрических токов возбуждения будут одни и те же в обоих случаях отведения. (Кроме того, при раздражении заднего корешка впереди узла токи возбуждения появляются в нервном стволе позади узла; следовательно, возбуждение в этом узле проводится в обоих направлениях точно так же, как и во всяком периферическом нервном волокне (Беритов, 4).
Характер рефлекса при раздражении чувствительного корешка. Роль узла в координации рефлекса можно исследовать, если вызывать рефлекс непосредственным раздражением самого чувствительного корешка. В некоторых случаях чувствительные волокна от одного рецептивного поля проходят через строго определенные задние корешки.
Если такой корешок будет перерезан, а его центральный конец будет раздражаться, то характер и координация вызванного рефлекса будут такими же, как и при раздражении самого поля. Например, у лягушки IX и X корешки получают волокна главным образом с рецептивного поля сгибания. Раздражение их вместе или в отдельности вызывает рефлекс сгибания (Беритов, 1). То же самое может быть показано и в отношении других рефлексов; например, Шеррингтои (2) показал это по отношению к рефлексу чесания.
В некоторых случаях, однако, раздражением чувствительного нерва не удается вызвать такой же эффект, какой получается при раздражении рецептивного поля. Нервные стволы и задние корешки большей частью содержат чувствительные волокна с рецептивных полей разных рефлексов. Поэтому раздражение их влечет за собой появление нескольких рефлексов в одном и том же двигательном органе. Следовательно, вызванное раздражением нервных стволов и задних корешков движение должно быть более сложным, чем от раздражения одного какого-либо рецептивного поля.
Такую картину очень часто можно видеть на лягушечьем препарате. Чувствительные нервы с рецептивного поля рефлекса потирания на бедре обыкновенно входят в мозг через VIII корешок, но нередко они частично входят и с IX корешком. Через IX корешок в мозг входят чувствительные волокна и из рецептивного поля сгибания. Поэтому очень часто раздражение IX корешка не вызывает исключительно одного какого-либо рефлекса.
Наоборот, последний представляет смешанное движение, которое включает элементы рефлекса сгибания и потирания: конечность сгибается, а пальцы производят ритмические движения, характерные для рефлекса потирания (Беритов, 3).
Таким образом, спинномозговые узлы, а также узлы черепно-мозговых нервов не принимают участия в координации рефлекса. Следовательно, центральная нервная система, г. е. спинной и головной мозг, представляет тот единственный орган, в котором протекают процессы координации рефлекса.
Сегментальное расположение координирующих аппаратов
Существование каждого рефлекса предполагает наличие известного рода аппаратов координации в спинном или головном мозгу. Еще Спалланцани (1778) ясно показал, что у жабы защитный рефлекс передних конечностей можно наблюдать и в том случае, если нервы передних конечностей находятся в связи с одним только III сегментам, т. е. когда спинной мозг перерезан впереди и позади данного сегмента. Отсюда видно, что координирующий аппарат защитного рефлекса должен находиться в III сегменте.
Ринберк (6) повторил этот опыт и на других животных. Он нашел, например, что у собаки для вызова грудных сегментальных рефлексов вполне достаточно наличия одного из грудных сегментов. Затем Данилевский (7), Бабак (8) и другие обнаружили, что поясничная часть спинного мозга лягушки способна на производство ритмических рефлексов, как ходьба и прыгание.
Все они ставили опыты с перерезкой спинного мозга между поясничной и грудной областями. Следовательно, координирующий аппарат ритмических рефлексов задних конечностей должен находиться в спинном мозгу, в его поясничном отделе. Такое же заключение было выведено относительно ритмических рефлексов у разных высших животных.
Для решения вопроса о местоположении координирующих аппаратов самым лучшим методом должно считаться локальное (местное) отравление центральной нервной системы стрихнином. Если отравить маленькой дозой стрихнина центральную нервную систему смазать 0.1 % раствором поверхность мозга или приложить к ней смоченную в этом растворе фильтровальную бумагу величиной в 1—2 мм2), то яд подействует на мозг характерным образом. Если смазать стрихнином заднюю поверхность спинного мозга лягушки, то это повлечет за собой необыкновенное усиление рефлекторных реакций, которые вызываются раздражением задних корешков отравленной части мозга (Беритов, 9).
Такое локальное отравление IX—X сегментов спинного мозга у лягушки способствует усилению тех оборонительных и ритмических рефлексов, которые обычно вызываются раздражением чувствительных корешков этих сегментов. Отравление же VII—VIII сегментов с задней поверхности усиливает тот рефлекс потирания, который обыкновенно вызывается с того рецептивного поля, где начинаются нервные волокна VII—VIII задних корешков (Дюссер де Барени, 10; Беритов, 9).
В то же самое время все другие рефлексы с других рецептивных полей протекают нормально. Так, например, на рис. 21 дается прекрасная иллюстрация того, как после стрихнинного отравления IX и X сегментов в одной симметричной половине мозга раздражение X заднего корешка отравленной стороны производит сильный стрихнинный эффект на задней ноге той же стороны, а такое же раздражение того же корешка другой стороны вызывает на той же ноге нормальный эффект без признаков стрихнинного отравления.
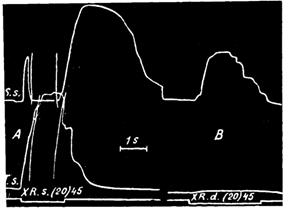
Рис. 21. Рефлекторные реакции из отравленного стрихнином и неотравленного участков мозга. Спинномозговая лягушка. Поясничный отдел мозга обнажен, и все задние корешки этого отдела перерезаны. В области IX—X сегментов с левой стороны на заднюю поверхность приложен стрихнин. Опыт записан в стадии сильного отравления. Верхняя кривая от m. semitend. sin., нижняя — от triceps sin. В опыте А раздражается X задний корешок отравленной стороны, при этом наступает сильный стрихнинный трехфазный эффект.
В опыте В раздражается X задний корешок правой стороны. В этом случае на той же конечности получается слабый сгибательный рефлекс. Внизу сигнальная линия отмечает время раздражения. На ней цифра в скобках обозначает ритм раздражения в 1 сек., а без скобок-силу раздражения в сантиметрах расстояния катушек
Вышеприведенные факты ясно указывают, что координация каждого рефлекса происходит как раз в том участке мозга, куда входят чувствительные нервы соответствующего рецептивного поля.
Принцип общего конечного пути. Изучение рефлексов показало, что начальный нейрон каждого рефлекторного пути, т. е. афферентный, принадлежит одной какой-либо рецептивной точке, в то время как конечный, эфферентный путь является общим для всех рефлекторных путей. Он является общим по отношению к рецепторам всего тела. Эта конвергенция или схождение всякого рода нервных путей афферентного, чувствительного характера в общем конечном двигательном пути происходит при помощи промежуточных нейронов, общих для определенной группы афферентных нейронов (Шеррингтон, 15).
Общие конечные пути однородны по своей функции: все они проводят возбуждение из центров к рабочему органу, причем они способны проводить возбуждение разных ритмов, вплоть до предельного ритма, характерного для двигательных нервов; когда производится два или больше раздражений в рецептивном поле одного рефлекса или вызывается два однородных или гармоничных рефлекса, то общий путь проводит возбуждение одновременно от рефлекторных путей того и другого рефлекса. Когда же происходит возбуждение рецептивных полей двух противоположных рефлексов, общий путь проводит возбуждение не от обоих рефлексов одновременно, а только от одного. В таких случаях говорят об антагонистических рефлексах.
Дата добавления: 2023-04-03; просмотров: 1280;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
