Понятия обратимых и необратимых процессов 4 глава
Самосборка глобулы характеризуется двумя основными путями. Это формирование плоской β-структуры, с последующим прилипанием к ней α-спирали, и, наконец, формирование β-«шпильки», с последующим изломом.
Доказано, что одному и тому же пространственному строению белка могут отвечать различающиеся последовательности аминокислотных остатков. Таким образом, можно предположить, что биологическая физика белка в какой- то степени определяется его пространственным строением, а α-спирали и β-формы являются вторичными структурами белка.
Доказано, что α-спирали и β-формы представлены в белке лишь частично. Они перемежаются неупорядоченными участками, благодаря которым белковая макромолекула сворачивается в глобулу, приобретая определенную пространственную, третичную структуру. В некоторых случаях белок обладает четвертичной структурой. В данном случае белковая система состоит из ряда глобул. Так, молекула гемоглобина состоит из четырех глобул, а белковая оболочка вируса табачной мозаики - из 2000 идентичных глобул.
Таким образом, белок – это многоуровневая система. Характер структуры ее на каждом уровне организации определяется геометрическими свойствами структур предыдущего уровня, силами взаимодействия их элементов и взаимодействия этих элементов с окружающей средой.
Доказано, что глобула белка формируется слабыми силами – электростатическими, Ван-дер-Ваальсовыми, а также водородными связями и, прежде всего, гидрофобными взаимодействиями. Глобулярные белки функционируют только в водном окружении. Вода влияет на водородную связь. Молекулы белка содержат как полярные, так и неполярные группы и располагаются таким образом, что первые группы контактируют с водой, а вторые удаляются из водного окружения белка. Среди аминокислотных остатков, из которых состоит белок, имеются полярные, неполярные и гидрофобные. Предполагают, что центральная часть глобулы белка - ее сердцевины должна быть гидрофобной («жирной»), а внешняя поверхность – гидрофильной («мыльной»). Водное окружение белка стабилизирует его электростатические солевые связи. Взаимодействие воды на солевую связь отличается от гидрофобного: солевые связи усиливаются, а гидрофобные ослабляются при добавлении неводных растворов.
Опыты показывают, что возможны фазовые переходы статического клубка в глобулу и обратно, то есть глобула является конденсированной кристаллической системой. Фазовый переход «белковая глобула – клубок» является переходом первого рода. Этот переход происходит при денатурации белка. Экспериментальное и теоретическое изучение денатурации белков дает информацию о степени стабильности белка (изменение его свободной энергии), таким образом, денатурация белков дает информацию об устойчивости внутренних связей, об электронно-конформационных взаимосвязях.
Существует корреляция между денатураций белков и температурой белка организма. Чем ниже температура тела, тем слабее внутримолекулярные взаимодействия, ответственные за жесткость молекулы, и тем ниже должна быть температура денатурации. Здесь особо важную роль играют междоменные взаимодействия в глобуле и электростатическое взаимодействие в солевых мостиках. С денатурацией белка хорошо коррелирует протеолиз, то есть гидролитическое расщепление белка осуществляют с помощью протеолитических ферментов; чем выше термостабильность белка, тем труднее он расщепляется.
3.2. Нуклеиновые кислоты
Нуклеиновые кислоты состоят из чередующихся звеньев фосфатной кислоты и сахара – углевода рибозы в рибонуклеиновой кислоте (РНК) и дезоксирибозы в дезоксирибонуклеиновой кислоте (ДНК).
К каждому углеводному звену присоединено одно из четырех азотистых оснований. ДНК и РНК – это тесты, написанные четырехбуквенным алфавитом. Они ответственны за биосинтез белков, за сборку их первичных структур.
ДНК – вещество генов, которое содержится в хромосомах и митохондриях клеток, а также бактериофагах. Молекулярные массы ДНК достигают 109. Это самые большие из известных молекул.
РНК фигурирует в различных формах, как в ядре, так и в цитоплазме клеток, а также в вирусах и фагах. Существует четыре типа РНК:
1) высокомолекулярные рибосомные РНК (рРНК) - с молекулярной массой порядка 106;
2) матричные, или информационные (мРНК) - с молекулярной массой 30000 и выше;
3) транспортные (тРНК) - с молекулярной массой 20000. Они содержат около 80 нуклеотидов;
4) высокомолекулярные вирусные РНК.
Существуют первичные и вторичные структуры в РНК и ДНК.
В настоящее время хорошо изучены структуры различных тРНК, известны “тексты” различных генов. Гены - это фрагменты (отрезки) молекул ДНК и РНК, которые программируют синтез белков, то есть являются ответственными за него. Таким образом, ген– это участок молекулы ДНК, в котором находится информация о первичной структуре какого-либо одного белка (один ген – один белок). Некоторые гены уже синтезированы. Ежегодно сообщается о синтезе все новых и новых генов.
Вторичная структура ДНК была установлена методом рентгеноструктурного анализа. Структура белка оказалась спиральной.
Информативная ДНК построена в виде двойной спирали, которая состоит из двух взаимно перевитых полинуклеиновых цепей, азотистые основания которых попарно соединены водородными связями. Обе цепи ДНК в двойной спирали взаимно камплементарны, то есть наблюдается однозначное соответствие между их нуклеотидами.
Доказано, что ДНК может кристаллизоваться в различных двухспиральных формах. При относительно низкой влажности ДНК кристаллизуется в так называемую α-форму. При большой влажности реализуется гексагональная β-форма.
Обе формы различаются значениями шага двойной спирали, углами поворота между соседними парами оснований и наклоном плоских пар к оси спирали. Если смотреть вдоль связи, вокруг которой происходит вращение спирали, то дальняя связь вращается по часовой стрелке относительно ближней. Эксперименты показывают, что все упомянутые двойные спирали нуклеиновых кислот имеют правое вращение, и только недавно было доказано, что существуют левые двойные спирали.
Специфическая двухсторонняя структура ДНК непосредственно объясняет ряд очень важных фактов – репликацию при митозе клеток и метаболическую устойчивость ДНК. При репликации двойная спираль сначала разделяется вследствие водородных связей и раскручивания цепей. Каждая цепь служит матрицей для сборки новой цепи, комплиментарной к матрице. Мономеры новой цепи соединяются с матрицей, образуя соответствующие пары. Одновременно происходит поликонденсация нуклеазитрифосфатов, и в результате образуются две двойные спирали, тождественные первичной. Такая полуконсервативная модель подтверждена экспериментами, содержит одну старую и одну новую цепь.
Из фага Т2 были выделены кольцевые двухспиральные молекулы ДНК длиной до 49 мкм. ДНК свернута в очень рыхлый клубок. Плотность ДНК приблизительно равна 1,7 г/см3. При нагревании и изменении рН происходит денатурация ДНК - переход двойной спирали в два клубка.
Круговой дихронизм (КД) ориентированных и неориентированных пленок ДНК меняется при изменении влажности и ионной силы.
Доказано, что структура двойной спирали ДНК и двухспиральных участков РНК определяется слабыми взаимодействиями – водородными связями, электростатическими и дисперсионными силами. Водородные связи возникают между различными атомами азотистых оснований ДНК. При плавлении двойной спирали водородистые связи между основаниями заменяются связями с молекулами воды. Азотистые основания спирали ДНК представляют собой π-электронные циклы, расположенные параллельно друг другу. Теоретические расчеты взаимодействий между азотистыми основаниями в ДНК показали, что Ван-дер-Ваальсовы силы в некоторых комплиментарных парах азотистых оснований значительно выше водородных взаимодействий.
В настоящее время разработаны методы расчета энергии горизонтальных и вертикальных взаимодействий атомов. Определяется сумма энергий электростатических и поляризационных взаимодействий и энергии отталкивания.
Денатурация нуклеиновых кислот сводится к разрушению двойной спирали ДНК или двухспиральных участков РНК. Нагревание раствора ДНК вызывает разделение двойной спирали на две цепи – сворачивающуюся и статического клубка. При этом уменьшается вязкость и оптическая активность. Возникает интенсивность поглощения света области 260 нм.
Доказано, что температура плавления ДНК растет с увеличением ионной силы раствора и пропорциональна логарифму концентрации катионов. Это объясняется тем, что чем выше концентрация катионов, тем в большей степени компенсированы отрицательные заряды фосфатных групп, следовательно, тем меньше отталкивание между комплиментарными цепями.
ДНК – уникальный пример одномерной упорядоченной структуры с апериодическим распределением звеньев. Термодинамические характеристики плавления ДНК были изучены методами микроколориметрии. Было доказано, что температура плавления сильно зависит от рН. При возрастании рН от 7,0 до 9,7 температура плавления убывает от 84,8 до 66,3 ºС. значение температуры плавления во многом зависит от ионной силы.
3.3. Биосинтез белка
Как известно, ДНК содержит информацию о первичной структуре белка, то есть генетическую информацию. Информация, записанная в первичной структуре белка, представляет собой последовательность нуклеидов.
Генетическая программа в вирусах РНК – тип программы, которая заложена в самих молекулах РНК. Основная проблема генетического кода заключается в выявлении соответствия между последовательностью аминокислотных остатков в белковой цепи.
Белковая цепь собирается непосредственно на двойной спирали ДНК, причем каждая аминокислота располагается в выемке между четырьмя нуклеидами, так как уже указывалось, что белковый текст ДНК (или РНК) – четырехбуквенный. Число нуклеидов, кодирующих один аминокислотный остаток, не может быть менее трех.
Доказано, что каждая выемка на двойной спирали имеет ромбическую форму. Два нуклеотида, как правило, принадлежат одной цепи, а следующих два – другой. Один из нуклеотидов первой цепи образует пару с нуклеотидами второй цепи.
Таким образом, составной код дает 20 «букв». Каждая «буква», то есть ромб, будет состоять из четырех нуклеотидов. Сочетание четыре по четыре, то есть 44=256.
Кодовое отношение было найдено экспериментально в результате генетических исследований, проведенных Ф.Х. Криком в 1961 г. Было доказано, что код универсален для всех организмов, начиная вирусами и кончая человеком. Код не может возникнуть в результате случайных встреч аминокислот. Он должен «набираться» на некоторой матрице.
матрицами являются молекулы ДНК и РНК. Для «набора текста» необходим генетический код. Матричная, или информационная, РНК переносит генетическую информацию от хромосом, в которых она находится, к рибосомам, на которых и реализуется биосинтез. Для биосинтеза необходим исходный материал – аминокислоты, которые фигурируют в клетке в свободном виде. Рибосомы представляют собой нуклеопротеидные частицы, которые состоят из белков и рибосомных РНК (рРНК).
Рибосомы обеспечивают надлежащее взаимодействие мРНК с тРНК, несущими аминокислоты, и поликонденсацию аминокислот в полипептидную цепь.
Как уже отмечалось, существует четыре типа РНК. В данном случае, при биосинтезе, информация, содержащаяся в генах, то есть в ДНК, перейдет в мРНК, то есть в так называемую «центральную догму» [ДНК->РНК->белок].
Вдоль одной цепи мРНК перемещается не одна, а ряд рибосом, на каждой из которых растет своя белковая цепь.
Система мРНК-рибосомы похожа на нитку с бусами (рис. 3.2) и называется полисомой. Синтез белковой цепи в полисоме есть перевод (трансляция) нуклеотидного текста в аминокислотный, происходящий с помощью генетического кода, то есть кодоаминокислотного словаря.
Таким образом, белковая цепь синтезируется из аминокислот, и далее из белковой цепи собирается биологически функциональная молекула белка.
Все сложные этапы биосинтеза характеризуются единым принципом, лежащим в основе молекулярной биофизики. Это принцип молекулярного узнавания, которое реализуется посредством слабых взаимодействий. В биосинтезе белков участвуют четыре типа ранее упомянутых нуклеиновых кислот: ДНК, мРНК, рРНК и тРНК. «Узнают» друг друга нуклеотиды комплиментарных цепей ДНК, нуклеотиды ДНК и мРНК, колонны мРНК и аминокислоты тРНК.
Узнавание реализуется и при взаимодействии нуклеиновых кислот с ферментами и рибосомами. В настоящее время рекомбинантные ДНК получают путем размножения двух спиралей ДНК. Этим занимается вновь возникшая наука - генная инженерия.
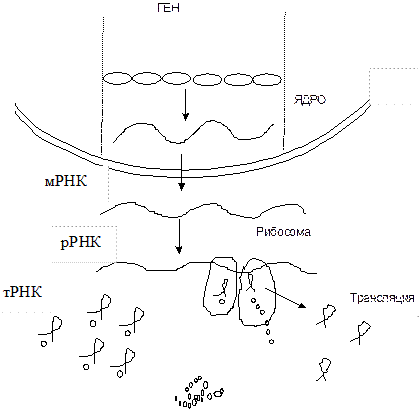
Рис. 3.2. Общая схема биосинтеза белка
Разрезание двойной спирали проводится с помощью специальных ферментов - рестриктаз. Полученный результат анализируется с помощью электрофореза в полиакриламидном геле. Такими методами генной инженерии определяют последовательность нуклеидов в ДНК, то есть структуры генов.
В настоящее время проводится синтез полных структурных генов и их внедрение в плазмоды или фаги, с помощью которых и размножаются молекулы ДНК. С помощью ДНК–транскритов осуществляется выделение генов.
Таким образом, генная инженерия обязана своим возникновением обратной транскрипции, которая позволяет изучить геномы вирусов, расшифровать молекулярные дефекты, лежащие в основе наследственных болезней крови, изучить молекулярные механизмы, ответственные за трансформацию нормальных клеток в опухолевые.
Трансляция – это перевод полинуклеотидного текста ДНК и мРНК в аминокислотный, белковый текст. Этот перевод происходит в комплексах рибосом с полирибонуклеотидными цепями, то есть в полисомах.
Структура рибосом изучается с помощью электронной микроскопии. Они представляют собой округлые гранулы диаметром 15…20 нм. В них находится РНК. Рибосомные РНК составляют примерно 65% сухого веса рибосом, белки – 35%.
Трансляция начинается с инициализации, то есть с синтеза первой пептидной связи. Далее происходит последовательная поликонденсация аминокислот. На протяжении всей трансляции растущий полипептид удерживается на рибосоме. Присоединение каждого следующего аминоацила идет на С-конце полипептида. Этот аминоацил пристраивается путем замещения тРНК на комплекс «аминоацил-тРНК». Этот процесс повторяется многократно в каждой рибосоме. Доказано, что в целом биосинтез белка определяется скоростями транскрипции и трансляции.
Первые опыты расшифровки генетического кода и его смысла были проведены Н.У. Ниренбергом (1961 г.). Он вводил синтетические полирибонуклеотиды в бесклеточную систему, которая содержала рибосомы, набор тРНК, АТФ и все необходимые ферменты, но не содержала ДНК и мРНК (система из разрушенных клеток). Исследователь наблюдал, что аминокислоты включаются в полипептидную цепь. Это было доказано путем центрифугирования и отмывания рибосом и тРНК. К смеси данных двух очищенных фракций добавлялась система, генерирующая АТФ-источник энергии. В бесклеточную систему вводились синтетические полирибонуклеотиды и изучалось включение меченых 14С аминокислот во фракцию, нерастворимую в полипептиде.
Таким бразом, оказалось возможным «обмануть» биологическую систему. Вместо природной (мРНК) рибосомы взаимодействовали с синтетическими полирибонуклеотидами.
В последующих опытах Н.У. Ниренберг использовал не полинуклеотиды, а тринуклеотиды известного строения. В данном случае в системе образовывались комплексы «тринуклеотиды – (тРНК) – аминокислоты» (аминоацил).
Были изучены все тРНК, которые последовательно связывались с мечеными аминокислотами. Так, были исследованы все 64 триплета и установлено, с какими аминокислотами они связаны.
Окончательную расшифровку, подтвердившую эти результаты, провел Х.Г. Корана (1966 г.). Им синтезировалась двойная спираль ДНК. Обе цепи спирали содержали комплиментарные, повторяющиеся n раз, триплеты. Причем каждая из цепей служила матрицей для транскрипции – синтеза полирибонуклеотида с помощью РНК – полимеразы. Таким образом, Х.Г. Корана получил две цепи, имитирующие мРНК с известной последовательностью повторяющихся триплетов. Обе цепи вводились в бесконечную систему, и определялось включение аминокислоты в полипептидную фракцию по методу Н.У. Ниренберга.
Эти опыты позволили проверить шесть кодонов (триплетов) в одном многостадийном синтезе. Полученный генетический код является универсальным. Основные кодоны действуют аналогичным образом в различных организмах. Универсальность кода доказывается размножением фагов и вирусов в клетках. Расшифровка генетического кода – это крупнейшее достижение молекулярной биологии и биофизики.
Таким образом, доказано, что для биосинтеза белка, для "набора текста" служат молекулы ДНК и РНК, то есть необходим генетический код. Матричный информационный принцип биосинтеза белка является основным для молекулярной биологии и молекулярной биофизики. Матричная, или информационная, РНК приносит генетическую информацию и реализует биосинтез. Для биосинтеза необходим исходный материал. Им служит аминокислота. Аминокислоты фиксируются в клетке в свободном виде. Биосинтез белка происходит на рибосомах. Последовательность событий биосинтеза белка является очень сложным, но все этапы характеризуются одним принципом, который лежит в основе молекулярной биофизики - это молекулярное узнавание, которое реализуется слабым взаимодействием. В биосинтезе участвуют четыре типа нуклеиновых кислот – ДНК, мРНК, рРНК, тРНК, которые узнают друг друга. Таким образом, нуклеиновые кислоты являются ответственными за биосинтез белков. В этом и состоит их так называемая «законодательная функция».
В свою очередь, «исполнительной властью» обладают белки - непрерывные участники всех процессов жизнедеятельности. Белки – ферменты, катализирующие все химические, электрохимические, механические процессы в клетках и организмах. Важнейшей функцией белка является ферментативная. Специализированные ферменты служат катализаторами всех метаболических реакций. Белки являются регуляторами генетических функций нуклеиновых кислот. Движение клеток и организмов, выполнение ими механической работы (например, мышечной) производится особыми сократительными белками, которые служат рабочими веществами этих процессов. Сократительные белки выполняют ферментативную АТФ-физную функцию, реализуют превращения химической энергии в механическую работу. Другие мембранные белки отвечают за транспорт молекул и ионов через мембраны клеток, за генерацию и распространение нервного импульса. Таким образом, белки определяют все метаболические и биоэнергетические процессы. Гамма-глобулины высших организмов защищают их от чужеродных биополимеров, функционируя в качестве антител в иммуных процессах. Итак, можно сказать, что белки являются обязательными участниками записания - передачи, трансформации и рецепции химических сигналов - макромолекул, молекул и ионов - в живых системах. Молекулы белков - самые сложные молекулы, известные науке. Так, глобулины - апериодические кристаллы сложной структуры. Наряду с глобулярными, существуют фибриллярные белки, сократимые и опорные. Большую роль играет денатурация белков - это утрата ими биохимической функции при нагревании.
В основные задачи физики белка входят:
1) теоретические и экспериментальные исследования структуры белковых молекул;
2) исследования, связанные между первичной структурой белковой цепи и пространственной структурой нативной макромолекулы;
3) изучение физических механизмов, лежащих в основе различных функций белков, прежде всего ферментной активности.
Вопросы для самопроверки
1. Перечислите виды «слабых» взаимодействий, существующих между белковыми субстанциями?
2. Что относится к «сильным» взаимодействиям структур надмолекулярных систем, содержащих белки?
3. Какова роль в организме гамма-глобулинов?
4. С помощью какого метода производят разделение белков?
5. Что называется денатурацией белков?
6. Что относится к первичной и вторичной структуре белка?
7. Из каких компонентов состоят нуклеиновые кислоты?
8. Перечислите четыре типа РНК?
9. Что называется геном?
10. В чем сущность биосинтеза белка?
11. Что называется трансляцией?
12. В чем заключается расшифровка генетического кода? Его смысл?
Тесты текущего контроля
1.1. Какие силы специфического отталкивания между полярными атомными группами определяют особенности структуры воды как конденсированной системы:
1) Индукционные силы; 4) Ориентационные силы;
2) Водородные связи; 5) Дисперсные силы или
3) Гидрофобные взаимодействия; силы Ван-дер-Ваальса.
1.2. Первичная структура белка – это своего рода текст, смысл которого состоит в биологическом функционировании белка, характеризующемся его структурой, написанной:
1) Пятибуквенным алфавитом;
2) Пятнадцатибуквенным алфавитом;
3) Двадцатибуквенным алфавитом.
1.3. Вторичными структурами белка являются:
1) 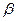 - шпильки; 3)
- шпильки; 3) 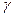 и - формы.
и - формы.
2) 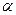 - спирали;
- спирали;
1.4. Из четырёх типов РНК найдите информационную РНК:
1) рРНК; 3) тРНК;
2) мРНК; 4) РНК.
1.5. Трансляция – это перевод полинуклеотидного текста ДНКи мРНК в аминокислотный белковый текст. Этот перевод происходит:
1) В аппарате Гольджи; 4) В метахондриях;
2) В полисомах; 5) В микротрубочках.
3) В комплексах рибосом;
ЛЕКЦИЯ 4. Физические свойства клеток
4.1. Строение и функции клеток и клеточных структур
Живая клетка - это органическая система, которая способна к самостоятельному существованию, развитию и воспроизведению. Клетка - основная биологическая структурная единица может существовать как отдельный организм. Примером являются бактерии, простейшие, некоторые водоросли. Известно, что одноклеточные организмы могут двигаться и защищаться от внешних воздействий. В них происходит обмен веществ. Наблюдается автономность клетки по отношению к окружающей её среде. Как известно, клетка состоит из цитоплазмы и внутриклеточного вещества, которое представляет собой сложный гель, в состав которого входят белки, углеводы, липоиды, органические кислоты, витамины и электролиты. Клетки обладают внешней мембраной, то есть оболочкой, отделяющей ее от окружающей среды, а также клетки имеют внутренние мембраны, отделяющие органеллы от цитоплазмы. Общая площадь мембран очень велика. Например, печень крысы имеет массу шесть грамм, а площадь мембран достигает сотни квадратных метров. Живая клетка – открытая термодинамическая система. В клетках синтезируются низко- и высокомолекулярные соединения, вплоть до белков и нуклеиновых кислот. Этот синтез происходит при участии ферментов, которые образуются внутри клеток. Кроме того, клетка запасает химическую энергию в молекулах (АТФ), за счет которой организм совершает механическую работу. Все эти процессы связаны с переносом веществ в клетку и выбросом из нее продуктов метаболизма.
Внутреннее содержимое клетки представлено цитоплазмой и расположенными в ней органоидами, или органеллами (рис. 4.1). Цитоплазма создает условия для осуществления физиологических реакций клетки и протекания биохимических процессов. Такое свойство цитоплазмы, как буферность, позволяет клетке осуществлять свою жизнедеятельность и поддерживать внутреннее постоянство среды при изменениях внешней, а постоянное движение - осуществлять связь между органоидами.

Рис. 4.1. строение клетки
4.2. Органеллы клеток
Примерно половина объема клетки занята органеллами. Органелла – специализированная субклеточная частица, выполняющая определенную функцию, например, ядро, эндоплазматический ретикулум (ЭР), аппарат Гольджи, лизосомы, митохондрии и др.
ЭР - система мембран, формирующих цистерны и каналы, разделяющие цитоплазму клетки на изолированные пространства или компартменты. Это необходимо, главным образом, для того, чтобы разделить множество параллельно идущих реакций. Мембраны ЭР служат местом протекания биосинтетических процессов. Выделяют шероховатый (Ш) и гладкий ЭР. Мембраны ШЭР содержат на своей поверхности рибосомы, на которых синтезируются белки, и представляют собой совокупность уплощенных мешочков. Строение гладкого ЭР ближе к трубчатому, и одна из основных его функций – синтез липидов. Помимо этого, каналы ЭР служат внутриклеточной системой переноса и распределения веществ. В зависимости от функционального состояния клетки мембраны ЭР подвергаются процессам сборки и разборки. Кроме того, эндоплазматическая сеть служит местом образования цистерн аппарата Гольджи.
Аппарат Гольджи (пластинчатый комплекс) представляет собой стопку из 5…30 уплощенных канальцев (цистерн) и связанных с ним пузырьков. Часто при описании трехмерной структуры аппарата Гольджи говорят, что он напоминает стопку блинов. У него выделяют наружную, обращенную к плазмалемме, и внутреннюю, соединенную с ЭР, поверхности. Функция данного органоида – транспорт и химическая модификация поступающих в него веществ. Кроме того, пластинчатый комплекс содержит собственные системы синтеза сложных углеводов из простых сахаров. Аппарат Гольджи представляет собой динамическую структуру, участвующую в потоке клеточных мембран. Он является промежуточным звеном между мембранами ЭР и плазмалеммой (наружная часть комплекса расходуется в процессе отшнуровывания пузырьков, а внутренняя постепенно формируется эндоплазматическим ретикулумом).
Рибосомы- мелкие органеллы, представленные глобулярными частицами диаметром порядка 20 нм, состоящими из двух субъединиц неравного размера – большой и малой. В состав рибосом входят белки и рибосомная РНК (рРНК), синтезируемая в ядрышке. Рибосомы могут либо свободно находиться в цитоплазме, либо прикрепляться к ЭР. На них происходит синтез белковых молекул (см. п.3.3.).
Митохондрии - в большинстве случаев палочковидной формы органоиды, размером несколько микрометров. Их содержимое – матрикс – окружено двумя мембранами. Внутренняя мембрана образует многочисленные гребневидные складки, называемые кристами (рис. 4.2). Митохондрии содержат мультиферментные системы, рибосомы и небольшое количество ДНК, чаще всего в виде кольцевых молекул. Митохондрии называют «энергетическими станциями» клетки, так как в них образуются молекулы АТФ, аккумулирующие энергию в виде химических связей. Митохондрии способны размножаться путем деления или отшнуровывания мелких фрагментов. Количество их в клетке зависит от функционального состояния и энергетических потребностей.
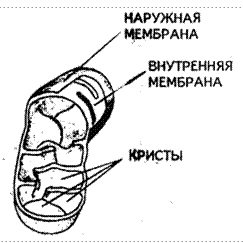
Рис. 4.2. Строение митохондрии
Лизосомы - мембранные пузырьки, содержащие литические ферменты гидролазы - протеазы, липазы, фосфотазы. Ферменты лизосом могут переваривать как поступившие в клетку путем эндоцитоза продукты, так и отдельные составные части клетки (а иногда ее целиком (автолиз). Лизосомы, отшнуровывающиеся от аппарата Гольджи, куда поступают ферменты, синтезированные в ЭР, называются первичными лизосомами. Они могут сливаться с пузырьками эндоцитоза или мембранами, окружающими ненужную структуру, образуя вторичные лизосомы, в которых происходит процесс переваривания и лизис содержащихся в них продуктов.
Пероксисомы, или микротельца, содержат фермент (каталазу), катализирующий разложение пероксида водорода на воду и кислород. Они окружены одинарной мембраной и имеют диаметр0,3…1,5 мкм.
Микротрубочки — трубчатые структуры цитоплазмы, состоящие из субъединиц глобулярного белка (тубулина), диаметром 10…25 нм. В клетке может происходить их сборка и разборка (например, при образовании веретена делений). Микротрубочки выполняют ряд функций, главными из которых являются: образование внутреннего каркаса, поддержание формы, образование нитей веретена деления. Кроме того, они могут служить направляющими при перемещении различных органелл, входить в состав ресничек и жгутиков.
Микрофиламенты - еще более тонкие, чем микротрубочки, нитевидные структуры (их диаметр около 7 нм), также состоящие из субъединиц белка (в основном актина) и подвергающиеся постоянной сборке-разборке. Микрофиламенты участвуют в создании тока цитоплазмы, в процессах движения клеток, а также эндо- и экзоцитозе.
Центриоли (у животных клеток) - мелкие полые цилиндры (длиной 0,4 мкм и около 0,2 мкм в диаметре), состоящие из 9 триплетов микротрубочек. В клетке два таких цилиндра, располагающихся перпендикулярно друг другу, лежат недалеко от ядра. При делении ядра эти структуры удваиваются и играют роль центров организации сборки нитей веретена.
Описание структуры, за исключением центриолей, характерно как для животных, так и для растительных клеток. Тем не менее между животными и растительными клетками существует ряд различий.
4.3. Строение ядра
От остальной цитоплазмы ядро отделено оболочкой, состоящей из двух слоев плазматической мембраны. Наружная мембрана переходит непосредственно в эндоплазматический ретикулум. Кариоплазма - внутреннее содержимое ядра - представлена гелеобразным матриксом, в котором располагаются хроматин и одно или несколько ядрышек. Хроматин представляет собой молекулы ДНК, связанные с белками. Он может находиться в виде тонких, неразличимых при световой микроскопии нитей (эухроматин), и в виде глыбок, лежащих, главным образом, по периферии ядра. В этом случае говорят о гетерохроматине. Различная степень конденсации (или спирализации) хроматина обусловлена разной генетической активностью расположенных в нем участков ДНК.
Дата добавления: 2020-11-18; просмотров: 455;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
