Понятия обратимых и необратимых процессов 7 глава

Рис. 4.24. Виды пассивного транспорта
Перенос через поры и эстафетную передачу веществ называют облегченной диффузией (см. рис 4.24).
Диффузией называется проникновение веществ без воздействия внешней силы. В данном случае процесс происходит без затраты энергии при перемещении вещества из места с большей концентрацией в место с меньшей концентрацией вследствие хаотического движения молекул.
Таким образом, пассивный транспорт осуществляется в результате диффузии веществ в сторону меньшего электрохимического потенциала.
Основным уравнением, характеризующим диффузию веществ через мембраны клеток, является уравнение Фика. Выведем уравнение Фика при рассмотрении переноса веществ в жидкостях.
Пусть через некоторую площадку S во всех направлениях перемещаются молекулы жидкости (рис. 4.25).
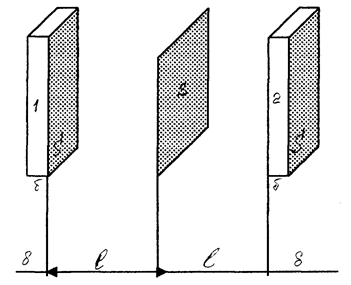
Рис.4.25. Проницаемость клеточных мембран (к выводу уравнения Фика)
Учитывая теорию молекулярного строения жидкости можно сказать, что молекулы пересекают площадку, перескакивая из одного положения равновесия в другое. Пусть l - среднее расстояние между молекулами жидкости; δ - толщина параллелепипеда (δ £ l); Sδ - объем каждого параллелепипеда.
Все молекулы, вследствие хаотичности их движения, можно условно представить шестью группами, каждая из которых перемещается вдоль или против направления одной из осей координат.
Так, в направлении, перпендикулярном площадке S, то есть вдоль оси ОХ, от первого параллелепипеда перескакивает 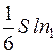 молекул, а противоположно оси ОХ -
молекул, а противоположно оси ОХ - 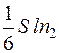 молекул, где n1 и n2 – концентрация молекул.
молекул, где n1 и n2 – концентрация молекул.
Исходя из рис. 4.25 и формулы (4.1), имеем:
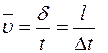 ,
,
где  - средняя скорость, м/с;
- средняя скорость, м/с;
Dt - время движения молекул, с.
отсюда
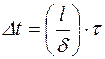 (4.27)
(4.27)
где τ- среднее время «оседлой жизни».
Тогда перенос молекул через площадку S за время 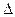 t будет
t будет
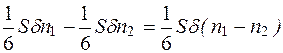 . (4.28)
. (4.28)
Умножая уравнение (7.2) на массу m отдельной молекулы и разделив на Δt, найдем поток вещества через площадку S:
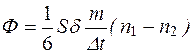 . (4.29)
. (4.29)
таким образом, получили количество вещества, которое за единицу времени проходит через площадку S.
Обозначим 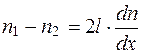 , тогда
, тогда 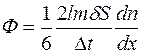 .
.
заменим 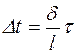 , тогда
, тогда
 . (4.30)
. (4.30)
Отношение потока к площадке, через которую он проходит, называется плотностью потока (кг/м3 или моль/м3).
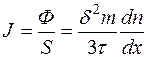 . (4.31)
. (4.31)
Известно, что массовая концентрация C=mn или n=C/m. Найдем
 , (4.32)
, (4.32)
тогда
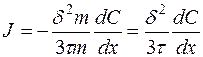 , (4.33)
, (4.33)
или, заменив 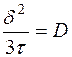 ,
,
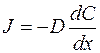 . (4.34)
. (4.34)
Получили уравнение Фика.
Знак минус показывает, что суммарная плотность потока вещества при диффузии направлена в сторону уменьшения концентрации, то есть в сторону, противоположную градиенту концентрации.
Коэффициент диффузии D зависит от природы вещества и от температуры. Природу характеризует τ - время релаксации, а расстояние δ между молекулами зависит от температуры Т.
Более простое уравнение диффузии предложено Коллендером и Берлундом:
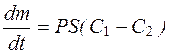 , (4.35)
, (4.35)
где С1 и С2 – концентрации вещества по разные стороны мембраны клетки;
Р – коэффициент проницаемости, аналогичный коэффициенту диффузии, но зависит не только от природы вещества и температуры, но и от свойств мембраны клетки и ее функционального состояния.
Если молекулы, диффундировавшие через мембрану вещества, движутся без образования комплекса с другими молекулами, то такая диффузия называется простой. Как отмечалось, кроме простой существуют облегченная и обменная диффузии.
При облегченной диффузии перенос веществ через мембрану происходит с помощью переносчиков и осуществляется с помощью каналов (рис. 4.26).
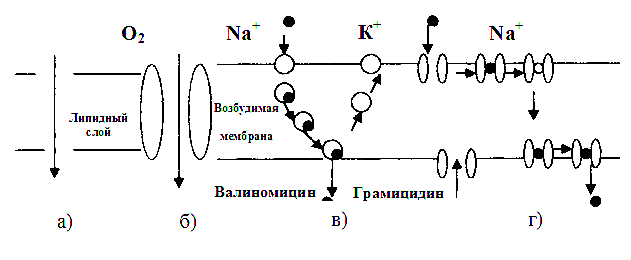
Рис. 4.26. Облегченная диффузия: а – простая диффузия; б – диффузия через канал; в – подвижный переносчик; г – эстафетный переносчик
Наличие переносчиков и каналов обусловливает селективность (избирательность) мембран.
В случае с простой диффузией считают, что избирательность связана с коэффициентом распределения К, а в случае облегченной диффузии - с избирательностью каналов и переносчиков.
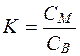 , (4.36)
, (4.36)
где СМ и СВ – концентрации веществ соответственно в масле и в воде.
Облегченная диффузия может осуществляться не только с помощью подвижных, но и с помощью фиксированных в мембране переносчиков.
Считают, что такие вещества, как глюкоза, глицерин, аминокислоты и ряд других, при определенных концентрациях проникают в клетки за счет облегченной диффузии, то есть посредством переносчика.
Наиболее подробно это явление изучено для случая переноса ионов ионофорными антибиотиками: валиномицином, грамицидином и другими, которые способны избирательно усиливать диффузию ионов через мембраны. Например, антибиотик валиномицин, образующий комплекс с ионами К+, повышает на несколько порядков проницаемость мембраны для калия.
Исследования механизма действия на мембрану подобных веществ представляют большой интерес для медицины, поскольку это открывает перспективы для создания новых эффективных анестетиков.
В настоящее время природа соединений, облегчающих диффузию, еще не установлена, хотя большинство ученых предполагают, что это – подвижные комплексы, растворимые в липидах клеточной мембраны. Этим объясняется проникновение в клетку веществ, нерастворимых в липидах и имеющих молекулы, большие по размерам, чем диаметр клеточных пор.
Таким образом, существуют два пути проникновения веществ в клетку через мембрану:
1) за счет растворения проникающих веществ в липидах клеточной мембраны (водонерастворимые органические соединения - органические жирные кислоты, эфиры);
2) через поры клеточной мембраны водорастворимых веществ и ионов, соединяющих цитоплазму клеток с внешней средой, к которым относятся: минеральные кислоты, соли и основания, сахара, спирты, мочевина и другие полярные органические соединения; то есть клеточная мембрана для неполярных соединений служит растворителем, а для полярных - молекулярным ситом.
Максимальной проникающей способностью через мембрану обладают вода и растворенные в ней газы.
Разновидностью облегченной диффузии является обменная диффузия (рис. 4.26, г). Сущность ее заключается в том, что вспомогательное вещество образует соединение с диффундируемым веществом (например, ион с молекулой лекарства) и перемещается к другой поверхности мембраны, где молекула проникающего вещества освобождается, на ее место присоединяется другая молекула этого же вещества и переносится обратно, образуя замкнутый круг. Концентрация по обе стороны мембраны не изменяется. Наличие обменной диффузии было доказано методом радиоактивных изотопов, используемых на эритроцитах.
Доказано, что натрий у эритроцитов, благодаря обменной диффузии, быстро обменивается на натрий плазмы, то есть одно вещество может обмениваться в эквивалентных количествах на другие вещества, находящиеся на противоположной стороне мембраны.
В последнее время получены ряд веществ, названных ионофорами, которые способны усиливать диффузию ионов через мембраны.
Перенос воды в клетках и тканях может осуществляться в результате осмоса, ультрафильтрации и электроосмоса.
Осмос - это движение молекул воды через полупроницаемую мембрану из области меньшей в область большей концентрации растворенного вещества.
Осмотическое давление (формула Вант-Гоффа):
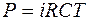 , (4.37)
, (4.37)
где i - изотонический коэффициент, который показывает, во сколько увеличивается количество растворенных частиц при диссоциации молекул неэлектролитов;
R - газовая постоянная;
С - концентрация.
Фильтрация – это перенос воды, который осуществляется при наличии гидростатического давления, то есть осуществляется с помощью формулы Пуазейля. С помощью фильтрации плазмы крови происходит образование первичной мочи в почечных нефронах.
Аномальный осмос – это перенос воды при одновременном наличии осмотического и электрического градиентов.
4.6.2. Активный транспорт веществ в клетках
Перенос молекул и ионов против электрохимического градиента за счет энергии гидролиза АТФ называется активным транспортом (рис. 4.27). Примерами активного транспорта могут служить перенос ионов калия и натрия через цитоплазматические мембраны (К+-, Na+-насос); - перенос Ca2+ через мембраны саркоплазматического ретикулума скелетных и сердечной мышц внутрь пузырьков ретикулума; перенос ионов водорода через мембраны митохондрий из матрикса наружу.
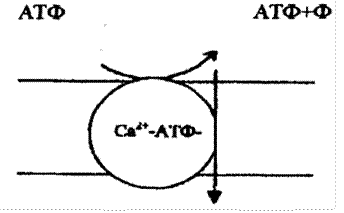
Рис. 4.27. Активный транспорт веществ
Наиболее сложно из транспортных АТФ устроена Н+-АТФ, состоящая из нескольких соединений. Самая простая – Са2+-АТФ, состоящая из одной субъединицы (полипептидной цепи), с молекулярной массой 100000. Рассмотрим механизм переноса ионов кальция этой АТФ.
Первый этап работы - связывание субстратов Са2+ и АТФ в комплексе с Mg2+. Эти два лиганда присоединяются к различным центрам на поверхности молекулы фермента, обращенной наружу пузырька саркоплазматического ретикулума (СР).
Второй этап работы фермента - гидролиз АТФ. При этом происходит образование энзимфосфатного комплекса (е-р).
Третий этап - переход центра связывания Cа2+ на другую сторону мембраны (транслокация).
Четвертый этап - высвобождение энергии макроэргической связи при гидролизе энзимфосфатного комплекса. Эта энергия расходуется на перенос Cа2+ с одной стороны мембраны на другую (внутрь пузырьков с меньшей концентрацией (1…4·10-2 M) в область с большей концентрацией (1…10·10-3 М)). Это и есть та работа, которую совершает Са2+ транспортная АТФ в мышечных клетках.
Для повторения цикла требуется возвращение кальцийсвязывающих центров изнутри наружу, то есть еще одно конформационное изменение в молекуле фермента. Молекулярный механизм натриевого насоса во многом сходен.
При этом клетка совершает работу, которая называется концентрационной, или осмотической. Эта работа состоит, с одной стороны, в накоплении клеткой веществ, присутствующих в окружающей среде в малых количествах, а с другой - в выведении в окружающую среду веществ, которые в ней присутствуют,имея высокую концентрацию. Величина концентрационной работы при переносе незаряженных частиц против концентрационного градиента может быть найдена из уравнения
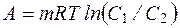 , (4.38)
, (4.38)
где m - количество молей вещества, перенесенного через мембрану из области большей в область меньшей концентрации.
Активный транспорт веществ можно разделить на два вида: 1) активный перенос ионов; 2) перенос органических веществ – главным образом, сахаров и аминокислот. К активному транспорту относят и такие явления, как пиноцитоз - так называемое «питье клеток», когда вещества поступают в клетку «упакованными» в клеточную мембрану.
4.6.3. Опыт Уссинга. Ионные каналы
Интересный пример активного транспорта представляет собой кожа лягушки. Если сосуд разделить на две камеры, содержащие нормальный раствор Рингера, свежеизолированной кожей лягушки, то внутренняя поверхность кожи заряжается положительно по отношению к наружной. Разность потенциалов при этом достигает примерно 100 мВ. Это так называемая «наносная» разность потенциалов, возникновение которой обусловлено активным переносом натрия. Компенсируя данную разность потенциалов с помощью внешнего источника, то есть ЭДС которого имеет противоположное направление, вновь наблюдается перенос ионов натрия от наружной поверхности кожи лягушки к внутренней. Этот перенос натрия, несомненно, связан с происходящим в коже обменом веществ.
Аналогичные явления наблюдаются в опытах с другими сложными мембранными системами. О высокой интенсивности процессов активного переноса ионов в клетках свидетельствуют данные термодинамических расчетов. Высчитано, что 70% энергии метаболизма мышцы лягушки в покое расходуются на перенос ионов натрия. Этот показатель возрастает на 50% в определенных условиях, когда стимулируется перенос ионов.
Следует отметить, что пассивный транспорт происходит по градиентам, которые, как правило, возникают в результате наличия активного транспорта. С другой стороны, сам активный транспорт - противоградиентный процесс. Он сопряжен с градиентным процессом более высокого порядка - гидролиза АТФ.
Таким образом, окислительные продукты питания, а затем гидролиз АТФ с освобождением свободной энергии и являются теми градиентными процессами, которые обусловливают противогра-диентные процессы в клетках и тканях, одним из которых является активный транспорт веществ.
Каким образом ионы переносятся через мембраны - с помощью специальных переносчиков или по каналам мембраны? Решение этого вопроса играет большую роль в объяснении передачи импульса по нервным волокнам. Опыты показали, что на 1 мкм2 мембраны находится несколько сот каналов.
Так, проводимость одиночного натриевого канала составляет 4·10 Ом-1, калиевого - 12·10-2 Ом. Пропускная способность первого ≈107 ион/с, второго ≈107 ион/с.
Ионный канал трактуют как своего рода «векторный фермент», катализирующий перенос иона. Активность этого «фермента» (то есть липопротеинового комплекса) регулируется электрическим полем.
Ионы, посредством каналов, являются источниками энергии, которая в дальнейшем тратится для распространяемого импульса. В основе всего процесса лежит ранее рассмотренный натриевый насос, который действует за счет энергии АТФ.
Ионные каналы высокоселективны, то есть сверхизбирательны.
Таким образом, клетки способны пропускать через свою оболочку самые различные вещества. Пассивный транспорт осуществляется в результате диффузии без затрат химической энергии. Основным уравнением, характеризующим диффузию веществ через мембраны клеток, является уравнение Фика.
Если перенос молекул и ионов осуществляется против химического градиента, то речь идет об активном транспорте. Этот перенос осуществляется за счет энергии метаболических процессов. При этом клетка совершает работу.
Вопросы для самопроверки
1. Где в клетке протекают биосинтетические процессы?
2. Что из себя представляет клеточная мембрана?
3. Приведите схемотическое изображение плазматической мембраны.
4. Какова роль липидов?
5. Из чего состоят липиды?
6. Перечислите органеллы клеток.
7. Перечислите основные функции клеток и клеточных мембран.
8. Чем определяется предельное разрешение оптического микроскопа?
9. Каковы недостатки электронной микроскопии?
10. В чем сущность метода «замораживание – скол – травление»?
11. Принцип рентгеноструктурного анализа.
12. Каков принцип работы поляризатора?
13. Сформулируйте закон Брюггера.
14. Запишите уравнение Малюса.
15. Чем определяется орбитальный магнитный момент?
16. В чем сущность эффекта Зеемана?
17. Чем определяется частота прецессии ядер и от чего она зависит?
18. Каковы виды пассивного транспорта? Приведите вывод уравнения Фика.
19. Активный транспорт веществ. В чем сущность калиево-натриевого насоса?
29. Виды искусственных мембран. Их строение.
Тесты текущего контроля
1. Жидкостно-мозаичная модель биологической мембраны включает в себя:
1. Белковый слой, полисахариды и поверхностные липиды;
2. Липидный монослой и холестерин;
3. Липидный бислой, белки, микрофиламенты.
2. Толщина биологической мембраны:
1. 10 нм
2. 0,1мкм
3. 10мкм
4. 10А°
3. Фазовый переход липидного бислоя мембран из жидко-кристаллического состояния в гель сопровождается:
1. Уменьшением мембраны;
2. Толщина мембраны не меняется;
3. Утолщение мембраны.
4. Липидная часть биологической мембраны находится в следующем физическом состоянии:
1. Жидком аморфном;
2. Жидкокристаллическом;
3. Твёрдом кристаллическом.
5.Удельная электроёмкость мембраны аксона:
1. 0,5·10-4 Ф/м2
2. 0,5·10-2 Ф/м2
3. 0,5·10-2 Ф/см2
4. 0,5·10-12 Ф/м2
задачи для самопроверки
1. При диффузии 5 мкг калия из аксонов кальмара во внеклеточную среду совершается работа 1,16 мДж. Определить концентрацию ионов калия в аксоне, если во внешней среде она равна 8 мм/л. Температура тела кальмара 10 ºС.
2. Для изучения структуры и функции биологических мембран используют модели – искусственные фосфолипидные мембраны, состоящие из биомолекулярного слоя – фосфолипидов. Толщина искусственной мембраны достигает около 6 нм. Найдите электроемкость 1 см2 такой мембраны, считая ее εr = 3. Сравните полученную электроемкость с аналогичной характеристикой конденсатора, расстояние между пластинами которого 1 мм.
ЛЕКЦИЯ 5. Электрические явления в клетках и тканях
Электрические явления в клетках и тканях играют огромную роль в жизнедеятельности организма. Они выполняют важнейшие физиологические процессы - возбуждают клетки и проводят возбуждение по клеткам и тканям.
Возникающие в живых клетках и тканях биопотенциалы тесно связаны с метаболическими процессами, происходящими в организме, они характеризуют физиологическое состояние организма и являются одним из чувствительных показателей происходящих изменений в клетках и тканях, норме и патологии.
5.1. Виды биопотенциалов. Их природа. Понятие двойного электрического слоя. Дзета-потенциал
Возникновение биопотенциалов связано с концетрационными градиентами, которые обусловлены несимметричным, неравномерным распределением ионов.
Хорошо теоретически разработана и экспериментально подтверждена мембранная теория Л. Бернштейна (1902 г.). В дальнейшем эта теория была развита в трудах М. Ходжкина, который в основу положил роль ионных градиентов в возникновении биопотенциалов и распределения их между клеткой и средой.
Были выдвинуты ряд других теорий, в частности фазовая, которая полностью не была подтверждена экспериментально. В последнее время на смену фазовой пришла полиэлектролитная теория. Согласно этой теории основой цитоплазмы служит полиэлекролитный гель сетчатой структуры с фиксированными на ней отрицательными зарядами.
При возбуждении полиэлектролитные структуры утрачивают избирательность, что вызывает диффузию натрия в клетку, а затем калия из клетки в окружающую среду.
Это является причиной деполяризации клетки, то есть измененияеё заряда. Все эти процессы способствуют возникновению биопотенциалов, к которым относятся:
1) диффузионные потенциалы;
2) фазовые потенциалы;
3) электродинамический и дзета-потенциал;
4) потенциал покоя;
5) мембранные потенциалы;
6) потенциал действия.
Диффузионные потенциалы возникают на границе раздела двух жидких сред в результате разлившей подвижности ионов. Их разность потенциалов находится из уравнения Гендерсона:
 , (5.1)
, (5.1)
гдеω - подвижность катиона, м2/(В·с);
V - подвижность аниона, м2/(В·с);
R=8,31 Дж/(моль·с) - газовая постоянная;
а1 - активность ионов в области, откуда идет диффузия;
а2 - активность ионов в области, куда идёт диффузия;
Т - абсолютна температура, К;
F=9600 Кл - число Фарадея.
Под активностью ионов понимают их концентрацию. Активность ионов всегда меньше их концентрации
a=fc, (5.2)
где f - коэффициент активности;
с - абсолютная концентрация.
Таким образом, диффузионная разность потенциалов зависит от разностей в подвижностях катиона и аниона и от отношения активностей ионов в измеряемых участках. При одинаковой подвижности катионов и анионов и при отсутствии концентрационного градиента диффузный потенциал Uд=0.
В биологических объектах диффузионный потенциал более отчетливо наблюдается при повреждении клеток.
Фазовые потенциалы возникают на границе раздела двух несмешивающихся фаз (например, раствор электролита в воде и какое-либо масло) в результате различной растворимости катионов и анионов в неводной фазе.
Поскольку цитоплазма клеток представляет собой многофазную микрогетерогенную систему, то на поверхности раздела фаз могут возникать фазовые потенциалы небольшой величины. Величину фазового потенциала можно определить из уравнения (5.1).
Электродинамический и дзета-потенциал. Поверхностный электрический заряд.Известно, что электрические заряды на поверхности коллоидных частиц или на поверхности клеток могут возникать в результате двух процессов:
1) диссоциации ионных групп;
2) адсорбции ионов дисперсионной среды на поверхности дисперсной фазы, которая сама не способна образовывать ионы.
Возникновение поверхностного заряда за счет ионизации может, например, происходить в белках и зависит от наличия кислотных и щелочных группировок, благодаря которым белки представляют собой биполярные ионы. В кислых растворах белок играет роль катиона.
В результате ионизации одни ионы (противоионы) уходят в дисперсную среду, а другие (потенциалообразующие ионы) остаются фиксированными на поверхности, содержащей данные молекулы белка. Поверхность, содержащая белки, будет иметь заряд того знака, который имеют потенциалообразующие фиксированные ионы, а дисперсионная среда будет иметь заряд, создаваемый противоионами. Такая, в целом нейтральная, система ионов называется двойным электрическим слоем.
Кровь - коллоидная система, и компенсация поверхностного электрического заряда форменных элементов должна осуществляться распределением ионов вокруг клеток крови, то есть последние имеют двойной электрический слой. Распределение противоионов в диффузной части двойного электрического слоя клеток крови зависит от природы их среды, концентрации электролита. Впервые теорию двойного электрического слоя взвешенных частиц разработал К. Гельмгольц. Двойной электрический слой, по К. Гельмгольцу, представляет собой конденсатор, обкладки которого состоят из ионов противоположного знака. Ионы, как одного, так и другого знака, прочно и неподвижно фиксированы, и поэтому двойной слой имеет постоянную структуру.
Представление Гельмгольца справедливо только для разбавленных растворов и представляет собой частный случай более общей теории, которая в дальнейшем была развита в трудах М. Гуи, О. Штерна (1936 г.), А.Н. Фрумкина (1952 г.) и др.
Гуи М. отметил, что противоионы не находятся в фиксированном состоянии, а пребывают в неупорядоченном тепловом движении. Помимо этого, противоионы подвергаются притяжению потенциалообразующимися ионами, находящимися на поверхности частицы. В результате взаимодействия этих двух сил - теплового движения и электрического притяжения - устанавливается равновесие, при котором плотность распределения ионов около поверхности частиц может убывать с увеличением расстояния от поверхности твердой фазы по закону распределения Больцмана. На структуру двойного электрического слоя коллоидных частиц оказывает влияние не только общая ионная сила раствора, но и другие свойства ионов суспензирующей среды, к которым следует отнести размер ионов, деформируемость их, степень гидратации, адсорбционную способность и др. Путем изменения рН среды или введения в систему поливалентных или легко деформируемых одновалентных ионов можно вызвать не только сжатие двойного электрического слоя частицы, но и изменить знак ее заряда.
Существует ряд мнений о природе электрического заряда клеток крови, но все они противоречивы. Кудряшов В.А считает, что электрический заряд на поверхности клеток и биосубстратов является следствием поверхностной адсорбции ионов. В результате ионизации противоионы уходят в дисперсионную среду, а потенциалообразующие ионы останутся фиксированными на поверхности дисперсной фазы клетки, обусловливая знак его потенциала. Однако Чижевский А.Л. доказал, что только 0,023% от всего количества отрицательных ионов плазмы принимает участие в образовании электрического заряда форменных элементов крови, а ионы натрия и калия являются ведущими в образовании их двойного электрического слоя и величины поверхностного потенциала.
Зееман Л. (1972 г.) считает, что основными факторами возникновения поверхностного электрического заряда клеток крови являются карбоксильные группировки нейраминовой кислоты. Поверхностный заряд клеток крови обусловливается степенью диссоциации кислотных или основных групп клеточной мембраны. В генезисе поверхностного потенциала принимают участие аминокислоты, а также фосфатные группировки фосфолипидов (С. Мериш, 1972 г.). Банган И. (1958 г.) полагает, что главная роль в определении эритроцитного заряда принадлежит кефалину, у которого остатки фосфорной кислоты обращены к наружной, а основные группы - ко внутренней стороне мембраны. Этим объясняется высокая плотность электрическою заряда эритроцитов. Аугер С. (1965 г.) показал, что снижение электрофоретической подвижности эритроцитов связано не с изменением поверхностной структуры эритроцитов, а с адсорбцией на их поверхности белковых молекул. В частности, С. Пироски с соавторами (1972 г.) считают, что эритроциты, нагруженные глобулинами, несут на себе более низкий электрический заряд. По расчетам С.С. Харамоненко, А.А. Ракитянской (1974 г.), максимальное количество адсорбированных на эритроците молекул равно примерно 1·10-1. Учитывая площадь эритроцитов, они делают вывод, что глобулины полностью экранируют находящиеся на эритроците элементарные электрические заряды. Вместе с тем, Н.И. Губанов считает, что заряд эритроцитов обусловлен диссоциацией кислотных групп молекул фосфолипидов (кефалина) на поверхности эритроцитов и не связан с процессами адсорбции белков и ионов. Приведенные данные позволяют сделать заключение, что происхождение заряда эритроцитов является многофакторным, и до настоящего времени нет удовлетворительного объяснения этого явления.
В создании поверхностного заряда тромбоцитов принимают участие фосфатные группировки фосфолипидов. Горин м. (1939 г.) считает, что клетку можно представить в виде заряженного электрошара, который при микроэлектрофорезе перемещается в однородном электрическом поле со скоростью
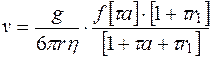 (5.3)
(5.3)
где r1 - радиус противоиона, Н·м;
 - вязкость среды, Па·с;
- вязкость среды, Па·с;
 - время, с.
- время, с.
A. Инер (1956 г.) при большом радиусе клетки и малой толщине ее двойного электрического слоя рассчитывают поверхностный электрический заряд по формуле
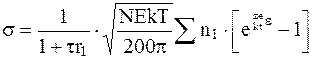 , (5.4)
, (5.4)
где k=5,67·10-8 Вт/(м2·К4) – постоянная Больцмана;
T – абсолютная температура, К;
Е - напряженность электрического поля, В/м.
Для симметричных электролитов уравнение (5.4) принимает вид:
 (5.5)
(5.5)
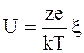 , (5.6)
, (5.6)
где 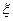 - дзета-потенциал, мВ;
- дзета-потенциал, мВ;
е=1,6·10-19, Кл – заряд электрона;
z - валентность.
Все данные уравнения связывают электрофоретическую подвижность форменных элементов крови с величиной их поверхностного заряда и с дзета-потенциалом. По мнению Э.П. Беликовой (1969 г.), дзета-потенциал - это потенциал между адсорбированными на поверхности клетки крови полиионами и их противоионной атмосферой.
Дата добавления: 2020-11-18; просмотров: 470;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
