Морфология крыльев и ног пчел: жилкование, структуры и таксономическое значение
Терминология жилкования крыльев. Крылья пчел проиллюстрированы, а их жилки обозначены на рисунке 10-6 с использованием модифицированной системы Комстока и Нидхэма. При описании крылья рассматриваются как расправленные, при этом направление к костальному краю (где расположена птеростигма на переднем крыле) называется передним, а направление к вершине крыла — дистальным. Для экономии места вместо полного термина "птеростигма" используется сокращение "стигма". Поскольку гомология жилок не всегда определена однозначно, а некоторые сходные по виду жилки, часто объединяемые в одну группу, имеют различные морфологические названия, предпочтительнее использовать термины, морфологически ни к чему не обязывающие при обозначении определенных ячеек и жилок. Такой подход широко применяется в таксономической практике.
Названия ячеек и некоторые нейтральные обозначения жилок показаны на рисунке 10-7, а в таблице 10-1 приведены эквиваленты этих названий в терминах системы Комстока-Нидхэма. Особое значение имеют три жилки, которые выглядят как поперечные: вторая абсцисса Rs, первая жилка r-m и вторая жилка r-m (по системе Комстока-Нидхэма). Именно эти структуры определяют количество субмаргинальных ячеек — обычно их три или две.
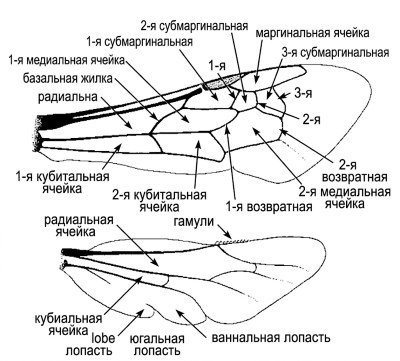
Рисунок 10-7. Схема крыльев пчелы, показывающая терминологию ячеек и морфологически нейтральные термины для обозначения определенных жилок. Обозначения 1, 2 и 3 относятся к субмаргинальным поперечным жилкам. Из работы Миченера, Макгинли и Данфорта, 1994.
Когда имеется только две субмаргинальные ячейки, иногда невозможно определить, какая именно жилка отсутствует — вторая абсцисса Rs или первая r-m. Как показал Питерс (1969), возможны оба варианта редукции, и каждый приводит к формированию двух субмаргинальных ячеек. Род Hyleoides (семейство Colletidae) (раздел 46) иллюстрирует эту проблему: невозможно точно установить, какая жилка утрачена. У представителей подсемейства Hylaeinae с двумя субмаргинальными ячейками первая обычно значительно длиннее второй, что позволяет предположить слияние первой и второй ячеек. Однако у Hyleoides наблюдается обратное соотношение, что затрудняет интерпретацию.
Упрощенная терминология поперечных жилок. Описание значительно упрощается при использовании сходной терминологии для всех трех жилок, которые в прошлом назывались первой, второй и третьей поперечными локтевыми венами, хотя эти структуры не имеют отношения к локтевой вене, расположенной в совершенно другой части крыла. Предпочтительнее обозначать их как первую, вторую и третью субмаргинальные поперечные жилки (1, 2, 3 на рис. 10-7). Эта терминология, предложенная Диницом (1963) по тем же соображениям, имеет лишь тот недостаток, что первая жилка технически не является поперечной, а представляет собой поперечный участок продольной жилки Rs.
Луис (1973) провел анализ предшествующей аларной терминологии и предложил новую номенклатуру жилок, пытаясь избежать вопросов гомологии и филогении. Он назвал субмаргинальные поперечные жилки первой-третьей RM, или радиомедиальными жилками. Однако менее техническое выражение "субмаргинальные поперечные жилки" дает более ясное представление о природе этих структур и их связи с субмаргинальными ячейками.
Методика измерений крыльевых структур. На рисунке 10-8 показаны стигма, маргинальная ячейка и прилегающие структуры; линии демонстрируют, как следует производить измерения, используемые в определительных ключах различными авторами. Ширина престигмы измеряется по костальному краю крыла, что терминологически не вполне корректно, поскольку полученная величина превышает ширину собственно престигмы. Длина костального края маргинальной ячейки, или длина участка ячейки, прилегающего к косте, используется для измерения расстояния от вершины стигмы до вершины ячейки.
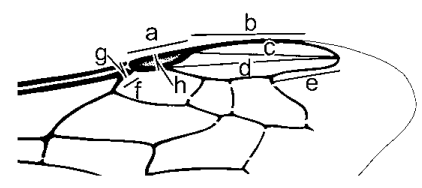
Рисунок 10-8. Переднее крыло Ceratina rupestris Holmberg с указанием способов измерений: (а) длина стигмы; (б) длина костального края маргинальной ячейки; (в) длина маргинальной ячейки за пределами стигмы; (г) общая длина маргинальной ячейки; (д) длина свободной части маргинальной ячейки; (е) длина престигмы; (ж) ширина престигмы (до края крыла); (з) ширина стигмы. Разрывы в жилках — боковые окна.
Скуловая доля и лопасть заднего крыла измеряются от основания крыла до вершин этих лопастей. На рисунках 10-6 и 10-7 видно, что длина скуловой доли составляет примерно две трети длины лопасти. Терминология, описывающая жилки и ячейки задних крыльев, варьирует: слово "брюшной" иногда заменяется на "анальный", а аббревиатура V — на А, поэтому брюшную долю иногда называют анальной долей.
Боковые окна представляют собой небольшие прозрачные участки, расположенные в определенных местах различных жилок (рис. 10-8). Линии сгибания, часто слабо различимые на перепонке крыла, пересекают жилки именно в этих окнах. Базальная жилка — общепринятый термин для обозначения первой абсциссы жилки М на переднем крыле (рис. 10-7).
Строение ног и ориентация структур. Некоторые авторитетные специалисты рекомендуют систему идентификации частей ног, предполагающую, что все ноги вытянуты в стороны под прямым углом к продольной оси тела. Несмотря на признание логичности такого подхода, здесь используется более традиционная система, при которой ноги рассматриваются в их естественном положении. Таким образом, бугорок голени находится на наружной, а не на передней поверхности задней голени, а два задних голенных отростка именуются наружным и внутренним, а не передним и задним. Дополнительные обозначения ориентации показаны цифрами на рисунке 10-9.
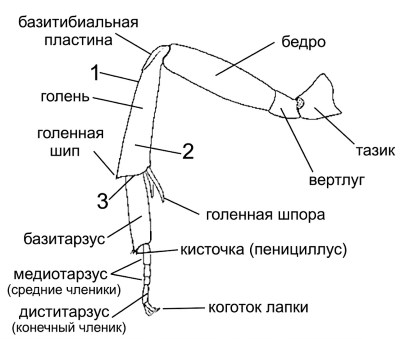
Рисунок 10-9. Задняя лапка самки пчелы с удаленными волосками, за исключением образующих пеницилл. Цифра 1 обозначает задний (верхний) край голени; 2 — наружную поверхность; 3 — дистальный (верхушечный) край. Модифицировано по Michener, McGinley and Danforth, 1994.
Голенные отростки (шпоры) представляют собой подвижные верхушечные выросты на голенях: один отросток (часть стригилиса) на передней голени, один на средней голени и, почти у всех пчел, два на задней голени. Голенные шипы (рис. 10-9) — это неподвижные, острые, расположенные дорсально апикальные выступы, обычно небольшого размера, часто тупые или мелкие. У некоторых пчел на голени имеется один, два или редко три шипа, расположенных непосредственно под углами.
Специализированные структуры ног. Чистительный аппарат передней ноги состоит из основной оси, или малуса, и тонкого пластинчатого скребка, или велюса, направленного к главной оси ноги; велюс обычно не достигает верхушки малуса. Шёнитцер (Schönitzer, 1986), а также Шёнитцер и Реннер (Schönitzer, Renner, 1980) ввели термин передний гребень для обозначения зубца или выступающего гребня на передней стороне голени у некоторых представителей надсемейства Apoidea.
Внутренняя поверхность задней голени имеет особое таксономическое значение. Обычно она несет два гребенчатых края, причем именно внутренний край часто модифицирован различными способами. Следуя традиции, этот край называется реснитчатым, если он несет тонкие, почти волосковидные выступы (обычно многочисленные). Во многих случаях такие выступы создают впечатление тонкого гребня. Мелкозубчатое или реснитчатое состояние (или промежуточные формы) характерно для перепончатокрылых (Gennerich, 1922). Аналогичным образом, наружная задняя голенная шпора и средняя голенная шпора часто описываются как простые, то есть немодифицированные.
Шпора называется гребенчатой, если ее внутренний край разделен на несколько длинных, грубых, часто тупых выступов, даже если в некоторых случаях их число сокращается до одного или двух. Форма задних голенных шпор и их связь с голенью различаются у разных таксонов и были подробно изучены Кейном (Cane, 1979).
Базитибиальная пластинка и щеточки. Базитибиальная пластинка (рис. 8-9, 10-9) расположена на верхней или наружной стороне основания задней голени многих пчел. Она лучше всего развита у самок и, предположительно, служит для опоры при передвижении пчел вверх или вниз по норкам в почве, а также при утрамбовке поверхностей ячеек пигидиальной пластинкой. Обычно базитибиальная пластинка окружена килем или иной четкой линией, и ее структура отличается от строения соседних областей. Однако пластинка может быть обозначена лишь рядом бугорков, как у многих Euryglossinae, или даже единственным бугорком, указывающим на ее вершину.
У некоторых пчел, например в определенных группах рода Centris, базальная или центральная часть базитибиальной пластинки резко приподнята над остальной поверхностью; эта область называется вторичной базитибиальной пластинкой. У большинства пчел с коротким язычком имеются парные щеточки или гребни, наиболее развитые у самок, на средних ногах. Одна щеточка расположена на нижней стороне голени (иногда также с щеточкой на базитарзусе), а другая — на базальной части бедра, иногда доходя до вертлуга.
Эти структуры расположены напротив друг друга и используются для очистки или переноса пыльцы с ипсилатеральной (той же стороны) передней ноги (Jander, 1976). Они называются средне-голенной щеткой (или гребнем) и средне-бедренной щеткой (или гребнем-щеткой). Гребень представляет собой один ряд щетинок, тогда как щеточка менее упорядочена.
Кейротрихии и структуры лапок. На внутренней поверхности задней голени большинства пчел имеется участок различного размера, покрытый волосками одинаковой длины, обычно тупыми, усеченными или слегка раздвоенными. Эти волоски, кейротрихии (рис. 10-11), по-видимому, служат для чистки крыльев. У некоторых пчел они замещены более длинными и обычными волосками, которые у самок могут выполнять функцию части скопы (собирательного аппарата).
На заднем базитарзусе многих самок пчел находится дистальный отросток, который выходит за пределы основания второго членика лапки (для простоты используется термин "членик лапки" вместо "тарсомер"). На вершине этого отростка иногда имеется небольшая кисточка — пенициллус (рис. 10-9), который не следует путать с пучком волосков на голени, известным как пенициллум у безжальных пчел (Meliponini).
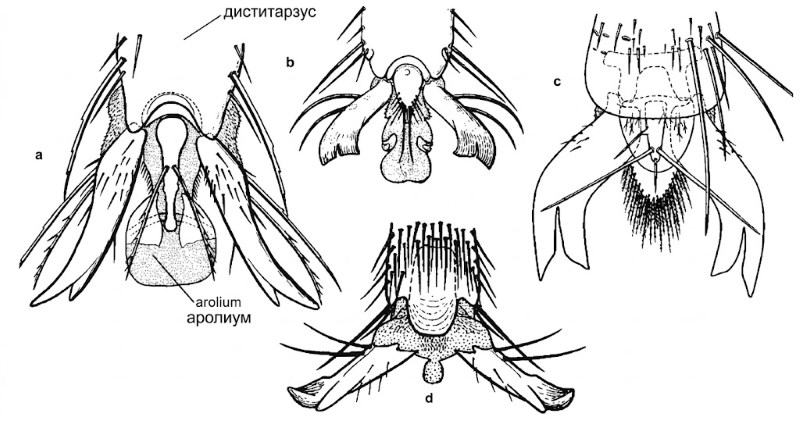
Рисунок 10-10. Виды верхушек лапок с обратной стороны: а — Andrena mimetica Cockerell; б — Anthophora edwardsii Cresson; в — Xylocopa orpifex Smith; г — Anthidium atripes Cresson. Аролии хорошо развиты на образцах а и б, отсутствуют или сильно редуцированы на образцах в и г. Срединная аролиевидная структура на образце г перепончатая, неразличима у сухих экземпляров; считается, что у Anthidium аролии отсутствуют. Из Michener, 1944.
Между коготками лапки часто имеется выступающий подушечкообразный аролиум (рис. 10-10). Его нижние дистальные поверхности почти всегда темные, часто черные, что помогает отличить его от сопутствующих бледных структур. Детальное строение структур между коготками описано Миченером (1944) и показано на рисунке 10-10. Вероятно, сравнительное изучение этих образований позволило бы выявить новые признаки, ценные для филогении и систематики пчел. Утрата аролиума у пчел происходила неоднократно в разных филогенетических линиях.
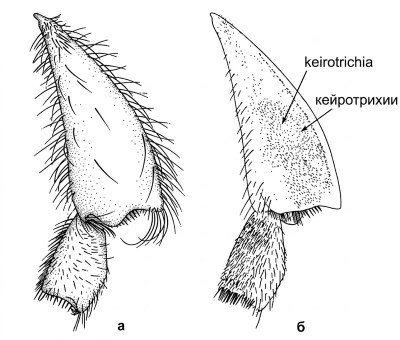
Рисунок 10-11. Задняя голень и базитарзус рабочей особи Plebeia frontalis (Friese): а — наружные поверхности, показывающие редукцию скопы до бахромы вокруг гладкой и преимущественно безволосой голени; б — внутренние поверхности. Из Michener, McGinley and Danforth, 1994.
Представленная здесь терминология и методики измерений имеют фундаментальное значение для таксономической практики. В последующих разделах систематической части эти признаки будут постоянно использоваться при описании родов и видов, что подчеркивает необходимость единообразного понимания морфологических структур. Детальное знание строения крыльев и ног позволяет не только точно идентифицировать таксоны, но и реконструировать филогенетические отношения, а также понимать функциональные адаптации, связанные с образом жизни различных групп пчел.
Сведения об авторах и источниках:
Авторы: Charles D. Michener
Источник: Пчелы всего мира
Данные публикации будут полезны студентам биологических специальностей, специализирующимся на энтомологии, аспирантам и исследователям в области эволюционной биологии, этологии и генетики насекомых, а также пчеловодам и всем, кто интересуется репродуктивными стратегиями и биологией развития общественных и одиночных пчел.
Дата добавления: 2026-03-15; просмотров: 311;
Поиск по сайту
Узнать еще
- ChatGPT для учителей: как использовать ИИ для непрерывного профессионального развития (НПР) и карьерного роста
- Автоматизация проектирования и система автоматизированного проектирования ПР
- Автосцепка АВ-6 Торецкого машиностроительного завода
- Автосцепка бывш. отделения промышленного транспорта ЦНИИ МПС
- Активные и пассивные операции Сберегательного банка России: структура и особенности
- Активные и пассивные операции центрального банка: структура и роль
- Аналитические исследования костного остова
- Анатомическая терминология и морфология пчел: строение головы и диагностические признаки

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
