Морфология хоботка и строение губ у пчел: терминология и диагностические признаки
Общая архитектура хоботка. Непропорционально большое количество признаков, используемых в систематике высшего уровня у пчел, базируется на особенностях строения хоботка, причем большинство склеритов этого сложного ротового аппарата хорошо различимы на рисунке 10-4. Однако терминология требует некоторых пояснений. В таксономической практике длину хоботка нередко выражают через точку, которой он достигает под телом пчелы при втягивании, то есть в сложенном состоянии покоя; это измерение имеет важное диагностическое значение при сравнении видов с разной трофической специализацией.
Хоботок пчелы морфологически подразделяется на три функциональных "сегмента". Базальный сегмент содержит кардо (cardo) — парные склериты максилл, которые в состоянии покоя направлены назад от мест сочленения с головной капсулой. Средний сегмент включает максиллярные щупики (palpi maxillares), предщупиковую часть галеа (pars praepalpalis galeae) и прементум (prementum); в покое этот сегмент выдвинут вперед от вершин кардо. Дистальный сегмент образован галеальными лопастями (lobi galeae), или постщупиковой частью галеа, лабиальными щупиками (palpi labiales), параглоссами (paraglossae) и глоссой (glossa); при сложенном хоботке этот сегмент вновь направлен назад, располагаясь над средним сегментом.
Структуры хоботка детально проиллюстрированы не только на рисунке 10-4, но также на рисунках 19-1 и 19-2, при этом терминология частей глоссы особенно обширна и представлена на рисунках 19-3 и 19-4. Важным элементом является галеальная пластинка — постщупиковая часть галеа, которая разделена продольным галеальным гребнем на две функциональные зоны: переднюю, тонкую, лишенную волосков, склеротизованные поверхности которой тесно сближены, и заднюю, несущую волоски и характеризующуюся значительным расхождением внутренней и наружной поверхностей. Передняя часть получила название галеальной оболочки (J. Plant, рукопись, 1991).
У пчел с коротким язычком (S-T пчелы) верхушки этих частей обычно разделены выемкой близ вершины галеальной пластинки, тогда как галеальный гребень, часто несущий ряд волосков, выступает в качестве укрепляющего элемента главным образом у пчел с длинным язычком (L-T пчелы). Предщупиковая часть галеа обозначается термином субгалеа (subgalea), что способствует прояснению сложной морфологии хоботка. Понимание этих структур критически важно для филогенетических реконструкций.
Склериты нижней губы: терминология и гомологии. В номенклатуре базальных склеритов лабиума существует значительная путаница. Основной склерит нижней губы, расположенный в среднем "сегменте" и служащий местом прикрепления лабиальных щупиков, обычно называют прементумом. У основания кардо и на мембране между верхушечными частями максилл располагаются один или два склерита, которые можно обозначить как лацинии (laciniae), хотя у большинства перепончатокрылых имеется только один такой склерит.
У многих пчел присутствует склерит, расположенный непосредственно у основания прементума, суживающийся к основанию и приобретающий более или менее треугольную форму; этот элемент обозначается как ментум (mentum) (Michener, 1944, 1985a), вслед за Снодграссом, который, однако, в 1956 году назвал его постментумом (postmentum). Плант и Паулюс (Plant and Paulus, 1987) считают, что это дистальная часть постментума, базальной частью которого является лорум (lorum). У некоторых пчел ментум частично (как у отдельных видов Andrena и Panurgus) или полностью (как у Ctenocolletes и некоторых Andrena) перепончатый (Michener, 1985a), но может быть идентифицирован по своему положению и форме.
У других пчел, например, у представителей семейства Halictidae, перепончатая область меньше, не имеет столь отчетливой формы и может представлять собой либо редуцированный ментум, либо просто мембрану между прементумом и постментумом. У большинства пчел базальным по отношению к лабиуму является лорум, который Миченер (1944) назвал субментумом (submentum), однако Уинстон (Winston, 1979) и другие исследователи отметили, что, поскольку у других перепончатокрылых имеется только один базальный губной склерит, лорум следует рассматривать как новую структуру, характерную исключительно для пчел и не гомологичную субментуму других отрядов насекомых.
Для удобства описания губные склериты, начиная с базальных, обозначаются как лорум, ментум и прементум. Когда лорум расширяется в виде слабой склеротизации, занимающей пространство между кардинами, эта структура именуется лоральным фартуком. В тех случаях, когда, как у большинства Halictidae, нижняя губа частично или полностью перепончата и ее склериты едва различимы, единственный базальный губной склерит предварительно рассматривается как лорум и лоральный фартук, а не как продукт слияния лорума и ментума.
Губно-челюстная трубка и связанные структуры. Базальная часть хоботка, непосредственно прикрепляющаяся к голове, называется губно-челюстной трубкой (обозначена пунктиром на рис. 21-2). Ее скелетными элементами являются кардо, представляющие собой прочные стержни в стенке трубки, дополнительно укрепленные гибкими полосами — конъюнктивальными утолщениями. Иллюстрации этих структур приведены на рисунках 21-2b и 59-1. У многих пчел нижний конец конъюнктивального утолщения обособлен в виде небольшого склерита — суспензория прементума, который соединяет утолщение с боковой выемкой в прементуме.
На задней поверхности губно-челюстной трубки ментум сильно варьирует, как описано выше и Миченером (1985a). Он может представлять собой довольно слабый плоский ментумный фартук — утолщение или склеротизацию задней стенки губно-челюстной трубки, занимающую большую часть пространства между кардинами. В других случаях ментум может быть более ограниченным по площади, но возвышенным вокруг соединения с прементумом, либо редуцированным до сильно склеротизованной V-образной структуры, соединенной медиально с прементумом (рис. 10-4а).
В двух последних вариантах, при втягивании нижней губы, основание лабиума и срединная (апикальная) часть ментума вместе обычно выступают, образуя выступ, отходящий кзади от губно-челюстной трубки. Эта структура называется хоботковой долей (рис. 21-2а). У некоторых пчел при втягивании хоботка данная доля вдвигается вверх и попадает в затылочный мешок — обширную ямку под затылочным отверстием (Roig-Alsina and Michener, 1993).
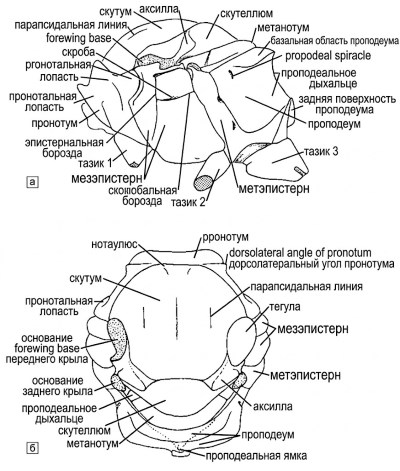
Рисунок 10-5. Схема грудной клетки пчелы: а — вид сбоку; б — вид сзади. На рисунке а опущена перегородка, на рисунке б — левая сторона. Проподеальный треугольник обозначен пунктирными линиями на рисунке б. Из работы Миченера, Макгинли и Данфорта, 1994.
Строение грудной клетки и ее склериты. Грудная клетка пчелы (рис. 10-5) представляет собой компактную структуру, образованную склеритами переднегруди (prothorax), среднегруди (mesothorax) и заднегруди (metathorax), к которым причленяются ноги и крылья, а также первого истинного брюшного сегмента, называемого проподеумом (propodeum). Переднегрудь представлена в основном крупной переднеспинкой (pronotum), которая простирается вентрально с каждой стороны в виде отростка, встречающегося или почти встречающегося со своим симметричным аналогом позади передних тазиков; проплевра (propleura) и простернум (prosternum) расположены перед этим латеровентральным расширением переднеспинки.
Переднеспинка служит важным ориентиром. Дорсолатеральный угол переднеспинки находится впереди и несколько латеральнее тегулы (tegula), часто с гребнем, килем или пластинкой, соединяющими эти структуры. Иногда между двумя дорсолатеральными углами вдоль заднего края переднеспинки проходит гребень, киль или приподнятая зона, называемая переднеспинным воротником; другой гребень или киль может проходить вертикально вниз от дорсолатерального угла к переднему тазику.
При рассмотрении сзади среднегрудь подразделяется на четыре основных склерита: щиток (scutum), щитик (scutellum) и парные аксиллы (axillae). Шов между каждой аксиллой и щитком называется аксиллярным швом; другие швы в этой области легко идентифицируются по названиям, например, щитково-щитиковый шов (scuto-scutellar suture). С боков среднегрудь представлена мезэпистернумом (mesepisternum), иногда называемым мезоплевроном (mesopleuron). Мезэпистернум может быть разделен почти вертикальной эпистернальной бороздкой (episternal groove), ранее именовавшейся предгрудной бороздкой.
Эпистернальная бороздка может проходить вниз, после соединения с передним концом горизонтальной скробальной бороздки (scrobal groove), в нижнюю переднюю часть мезэпистернума, как у большинства Colletinae и Halictinae (рис. 20-5b). В других случаях она может изгибаться кзади дугой, неразличимо сливаясь со скробальной бороздкой, как у Andrena и многих Halictinae (рис. 20-5c), либо полностью отсутствовать (рис. 20-5а), или быть настолько короткой, что не достигает скробальной бороздки, как у Megachilinae.
Скроб (scrobe) представляет собой небольшую ямку на скробальной бороздке перед мезо-метэпистернальным швом. Область над скробальной бороздкой и за эпистернальной бороздкой часто более выпуклая и блестящая, чем соседние участки; она называется гипоэпимеральной областью (не обозначена на рис. 10-5). Согласно Бразерсу (Brothers, 1975), вдавленный и в значительной степени скрытый передний край мезэпистернума именуется препектусом (prepectus).
Латеральная (в отличие от вентральной) часть мезэпистернума подразделяется на переднюю и боковую поверхности, причем угол между ними варьирует от плавно закругленного (рис. 20-5а) до резко заостренного и даже пластинчатого. Чтобы избежать громоздких описаний, Миченер и Гризволд (Michener, Griswold, 1994a) ввели термин омаулус (omaulus) для обозначения этого угла, расширив его использование по сравнению с терминологией сфекоидных ос, где он применяется только при наличии киля. Таким образом, можно лаконично описать: "омаулус округлый", "омаулус килевидный" или даже "омаулус пластинчатый".
Строение заднегруди и проподеума. В дорсальной части заднегрудь состоит из склерита метанотума (metanotum), который косо разделен с каждой стороны трансметанотальным швом. Метэпистернум (metepisternum), или метаплеврон, образует боковую поверхность заднегруди. Основания крыльев расположены над верхними краями мезэпистернума и метэпистернума. Средние и задние тазики пчел внешне смещены кзади, так что средняя нога кажется выходящей из нижнего конца метэпистернума, а задняя — из проподеума (рис. 10-5а).
Форма и подразделения проподеума имеют важное систематическое значение. У многих пчел на проподеуме имеется пара вдавленных линий (пунктир на рис. 10-5б), которые начинаются около его передних дорсолатеральных частей и простираются вниз и заднемедиально, почти сходясь в проподеальной ямке или над ней — срединном углублении на нижней части задней поверхности. Эти линии вместе с передним дорсальным краем проподеума образуют проподеальный треугольник, который морфологически представляет собой метапостнотум (metapostnotum) (Brothers, 1976).
Форма проподеума при рассмотрении сбоку совершенно независима от треугольника. Весь проподеум может быть вертикальным или почти вертикальным, спускаясь от заднего края метанотума (рис. 20-5а, в), — в этом случае он называется наклонным. Часто имеется более или менее горизонтальная или наклонная базальная область (рис. 20-5б), иногда отделенная резкой линией или килем от наклонной задней поверхности, как показано на рис. 10-5а. Горизонтальная часть называется базальной зоной или базальной областью проподеума, а иногда — ограждением (enclosure), когда она выделена или окаймлена килями.
Базальная область может быть частью проподеального треугольника или выходить за его пределы, по крайней мере латерально. Иногда базальная область, судя по ее форме, вертикальна, как и остальная часть задней проподеальной поверхности. Термин "базальная зона" применим даже в тех случаях, когда она не отделена от вертикальной поверхности четкой линией и даже когда она наклонная или вертикальная, а не горизонтальная. Однако у некоторых пчел обе поверхности непрерывно закруглены, переходя одна в другую в виде широкой изогнутой поверхности, что делает невозможным определение базальной зоны при отсутствии характерного рельефа.
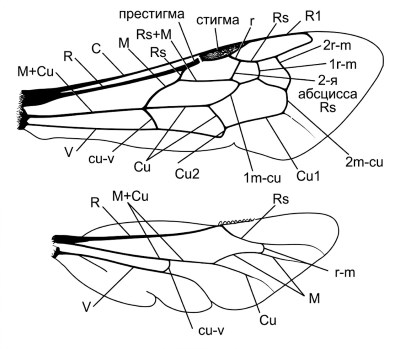
Рисунок 10-6. Схема крыльев пчелы, иллюстрирующая терминологию Миченера (Michener, 1944). По материалам Michener, McGinley, and Danforth, 1994.
Жилкование крыльев. Жилкование крыльев (рис. 10-6) предоставляет обширный набор диагностических признаков, широко используемых в систематике пчел на всех уровнях — от семейств до видов. Терминология жилок и ячеек следует системе, разработанной Миченером (1944) и основанной на гомологизации с общим планом строения крыла перепончатокрылых. Основные продольные жилки включают костальную (C), субкостальную (Sc), радиальную (R), медиальную (M), кубитальную (Cu) и анальную (A). Поперечные жилки соединяют продольные, ограничивая ячейки.
Наиболее важными для диагностики являются форма и размер радиальной ячейки, количество и форма субмаргинальных ячеек, наличие или отсутствие маргинальной ячейки, а также характер жилкования в вершинной части крыла. У представителей разных семейств наблюдаются характерные особенности жилкования: например, для Apidae типично наличие трех субмаргинальных ячеек, для многих Halictidae — двух, а у Megachilidae форма радиальной ячейки и субмаргинальных ячеек варьирует в широких пределах. Редукция жилкования характерна для некоторых паразитических групп, таких как Nomadinae, у которых отдельные жилки могут полностью исчезать.
В следующих разделах, посвященных систематическому обзору семейств и родов, терминология, изложенная выше, будет постоянно использоваться при описании диагностических признаков, что подчеркивает необходимость единообразного понимания морфологических структур. Представленные здесь иллюстрации и пояснения призваны служить справочным аппаратом при работе с таксономическими ключами и описаниями видов, обеспечивая корректную интерпретацию анатомических особенностей, значимых для идентификации и филогенетических построений.
Сведения об авторах и источниках:
Авторы: Charles D. Michener
Источник: Пчелы всего мира
Данные публикации будут полезны студентам биологических специальностей, специализирующимся на энтомологии, аспирантам и исследователям в области эволюционной биологии, этологии и генетики насекомых, а также пчеловодам и всем, кто интересуется репродуктивными стратегиями и биологией развития общественных и одиночных пчел.
Дата добавления: 2026-03-15; просмотров: 250;
Поиск по сайту
Узнать еще
- Анатомическая терминология и морфология пчел: строение головы и диагностические признаки
- Анатомическое строение масличных семян
- Анатомия птицы: систематическое положение, внешнее строение и особенности вскрытия сизого голубя
- Астеносфера Земли: строение, свойства и роль в глобальной геодинамике
- Бактериофаги. Описание и строение
- Блоки зубчатых колес. Машиностроение
- Вибрационное опыление и эволюция пчел: механизмы специализации и экологии
- Влияние отделки и плотности на строение и свойства тканей

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
