Скопальные структуры и перенос пыльцы у пчел: морфология собирательного аппарата
Строение и локализация скоп. У самок пчел имеются специализированные структуры для удержания и транспортировки пыльцы — скопы (scopae), тогда как у самцов они отсутствуют, за исключением представителей подсемейств Hylaeinae и Euryglossinae семейства Colletidae. Скопы также отсутствуют у пчел-паразитов и пчел-грабителей из различных семейств, а также у маток высокоэусоциальных пчел (Meliponini и Apini). В определительных ключах и описательных комментариях термин "скопа" используется без дополнительных напоминаний о том, что эти структуры встречаются только у самок. Важно понимать, что скопа образована волосками, предназначенными именно для транспортировки пыльцы в гнездо, а не для ее первичного сбора с цветков, который осуществляется другими щеточками и кисточками.
Некоторое количество пыльцы может переноситься на различных частях тела, но основные скопы локализованы главным образом на задних конечностях (рис. 6-4, 8-5) или на вентральной стороне метасомы (рис. 8-7). У большинства пчел скопальные волоски располагаются на задних лапках, однако у непаразитических представителей семейства Megachilidae они находятся на метасомальных стернитах. У некоторых Colletidae и Halictidae скопы присутствуют как на нижней стороне метасомы, так и на задних ногах, что представляет собой переходное состояние между разными типами локализации собирательного аппарата.
Если окаймления из скопальных волосков окружают пространство, в котором транспортируется пыльца, такие структуры называются корзиночками (corbicula). Наиболее известная корзиночка расположена на наружной стороне задней голени у представителей семейства Apidae (трибы Euglossini, Bombini, Meliponini, Apini). Она образована не только волосками, но и замкнутой вогнутой или плоской поверхностью голени (рис. 10-11). Другие типы корзиночек встречаются на нижней стороне задних бедер у Andrenidae, Halictidae, Colletidae и других семейств, а также по бокам проподеума у многих видов Andrena, некоторых Halictidae и Colletidae.
Метасомальная структура и сегментация. Для простоты и экономии места, как отмечено в разделе 9, метасомальные тергиты и стерниты обозначаются сокращенно: T1, T2 и т.д., а также S1, S2 и т.д., причем T1 и S1 образуют базальный сегмент метасомы (рис. 10-12). Каждый тергит или стернит (за исключением переднего и редуцированного верхушечного) состоит из основной пластинки, на которой обычно различимы несколько поперечных линий. По переднему краю, всегда полностью скрытому в интактной метасоме, проходит переднебоковой шов, с эквивалентным внутренним гребнем, служащим местом прикрепления продольных межсегментарных мышц.
Очень узкие края тергита и стернита перед этим швом называются акротергитом и акростернитом. От них отходит аподемальный край, который расширяется, образуя поперечные аподемы. Ближе к середине каждой пластинки проходит еще одна поперечная линия — градус (gradulus). Поверхность, расположенная базальнее градуса, т.е. преградусная область, обычно слегка приподнята по сравнению с постградусной, что делает градус заметным в виде невысокого валика, как показано на рис. 10-12b.
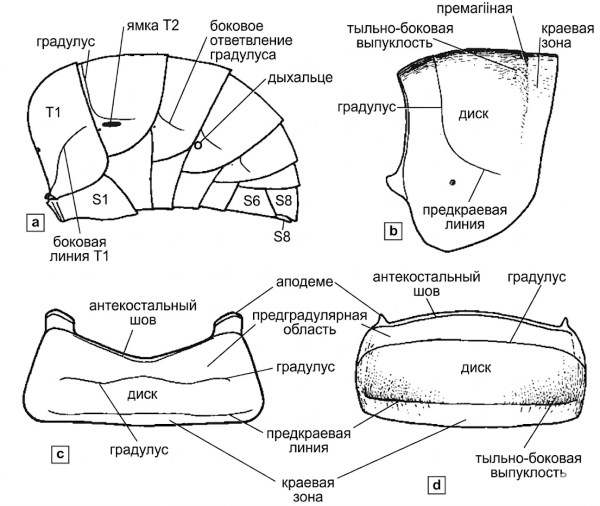
Рисунок 10-12. Метасомальные структуры самца пчелы: а — схематический вид метасомы сбоку; б — вид сбоку T3; в — вид спереди S3; г — вид сзади T3. а, в и с модифицированы по Michener, 1944.
Концы тергальных градусов, если они не загнуты резко назад, обычно располагаются вблизи дыхалец. Если же они сильно загнуты назад, образующиеся продольные линии называются боковыми частями градуса или боковыми ответвлениями градуса. В случае изгиба формируются боковые градусы, которые иногда приподняты, образуя боковые градусные пластинки или боковые градусные кили. Градусы — за исключением их боковых ответвлений — часто скрыты предшествующим тергитом или стернитом на неповрежденной метасоме, однако на T2 и S2 они иногда обнажены или могут быть легко обнажены при легком растяжении метасомы.
Вблизи заднего края каждого тергита и стернита обычно проходит еще одна поперечная линия — премаргинальная линия, отделяющая краевую зону (задняя краевая область по Миченеру, 1944; апикальное вдавление по Тимберлейку, 1980b) от остальной части склерита (рис. 10-12). Эта зона часто вдавлена, но в других случаях отличается только скульптурой от прилегающей области, а иногда краевая зона вообще не дифференцирована. Область между градусом и премаргинальной линией может называться диском, хотя иногда, как на T2 у Exomalopsis, премаргинальная линия выгибается далеко вперед, так что краевая зона становится широкой, а диск сужается до поперечной полосы.
Специализированные области тергитов. Дорсолатеральные части дисков (между градусом и премаргинальной линией), особенно на T2–T4, часто несколько приподняты, выпуклы и нередко блестящи. Эти дорсолатеральные выпуклости часто подчеркнуты премаргинальными линиями, которые ограничивают их сзади. T1 отличается от остальных тергитов тем, что его основание сужено из-за узкого соединения с грудным отделом, однако его дорсальная горизонтальная поверхность сходна с поверхностью последующих тергитов, часто имея краевую зону, премаргинальную линию и диск. Ближе к основанию T1 сильно наклонен, и часто вблизи вершины наклонной поверхности имеется поперечная линия, более или менее отделяющая ее от горизонтальной поверхности; это может быть градус T1, хотя данный термин для этой структуры обычно не используется.
Эта линия (или киль), ограничивающая переднюю поверхность или переднюю вогнутость T1, хорошо развита у представителей рода Heriades и некоторых других мелких Megachilidae. Кроме того, обычно присутствует боковая линия T1 (рис. 10-12а). У различных Colletidae и Andrenidae часто развита латеральная ямка T2 (рис. 10-12а), коррелирующая с наличием лицевых ямок. Предположительно, функция латеральных ямок T2 сходна с функцией лицевых ямок; у самцов обе ямки слабо выражены, но часто различимы у самок.
Тергиты часто несут поперечные полосы светлых волосков, которые могут располагаться на разных частях. Термины "метасомальные полосы" и "фасции" используются взаимозаменяемо: апикальные полосы находятся на краевых зонах или перекрывают их, а базальные полосы расположены на дисках. Эти структуры имеют важное диагностическое значение при идентификации видов.
Пигидиальная пластинка и связанные структуры. Пигидиальная пластинка (рис. 10-13) обычно представляет собой плоский склерит, окруженный с боков и сзади килем или отчетливой линией. У самок она расположена на T6, у самцов — на T7, иногда формируясь в виде апикального выступа. Линия, ограничивающая пигидиальную пластинку, может быть срединным углублением градуса T6 (у самок) или T7 (у самцов), как хорошо видно, например, у самок Eucerini. У многих пчел градус отсутствует латерально, так что присутствует только часть, ограничивающая пластинку.
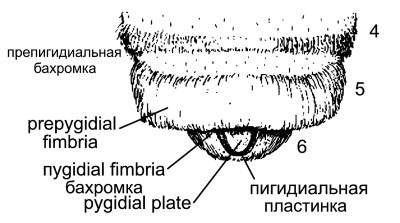
Рисунок 10-13. Схема верхушки метасомы самки пчелы, например эвцерины. Номера тергитов указаны с правой стороны.
В других случаях градус полностью поперечный, т.е. проходит через базальную часть тергита до пигидиальной пластинки, которая тем не менее остается ограниченной. Это наблюдение заставляет предположить, что пигидиальные пластинки могут иметь два негомологичных происхождения. Иногда пигидиальная пластинка редуцирована до плоского острия или полностью отсутствует; у самцов она отсутствует или рудиментарна чаще, чем у самок.
Препигидиальная фимбрия представляет собой полосу густых волосков на вершине T5 у самок, заметно отличающуюся от апикальных полос волос или фасций, которые могут присутствовать на предыдущих тергитах (рис. 10-13). Считается, что препигидиальная фимбрия отсутствует, если полоса волос на T5 сходна с таковой на T4 и предшествующих тергитах. У самок подсемейства Nomadinae препигидиальная фимбрия модифицирована и состоит из однородных коротких волосков на часто наклонной поверхности апикальной срединной части T5, образуя область, напоминающую пигидиальную пластинку, иногда отчетливо очерченную гребнем или изменением наклона и текстуры поверхности; эта структура называется псевдопигидиальной областью.
Густые волоски на T6 самок, расположенные с каждой стороны пигидиальной пластинки, образуют пигидиальную фимбрию (рис. 10-13), которая разделена пластинкой на две части. T7 самки, всегда полностью скрытый, состоит из двух слабо склеротизованных пластинок, называемых гемитергитами T7, которые являются частью жалового аппарата. Каждый из них содержит дыхальце, по которому их легко идентифицировать (рис. 10-14).
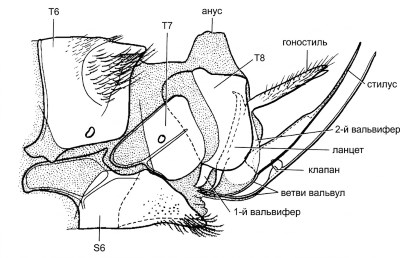
Рисунок 10-14. Верхушка метасомы самки Halictus farinosus Smith с искусственно выдвинутым жалом. Из Michener, 1944.
Гемитергиты T8 представляют собой пластинки сходного размера, лишенные дыхалец. Вторые клапаны, или женские гонококситы, также имеющие пластинки без дыхалец, дают начало базальным ветвям вторых клапанов и апикальным третьим клапанам, или женским гоностилям, которые также называются жалящими влагалищами. Другая терминология включает: стилет для сросшихся вторых клапанов, ланцеты для первых клапанов и клапан для дорсальной створки у основания каждого ланцета. Маленький первый клапан в литературе о медоносных пчелах называется треугольной пластинкой.
Гениталии и терминалии самцов. Гениталии, а также стерниты S7 и S8 самцов пчел обладают многими диагностическими признаками и могут быть извлечены для изучения, хотя у фиксированных экземпляров они почти всегда втянуты. У некоторых групп, таких как Heriades из семейства Megachilinae, передние стерниты, включая S6 и S5, и редко даже S4 и S3, также скрыты и модифицированы, однако термин "скрытые стерниты" обычно применяется только к S7 и S8.
У свежего или размягченного образца обычно можно проникнуть между верхушечными тергитами и стернитами и с помощью иглы с крючком извлечь гениталии и скрытые стерниты. В большинстве случаев такая препаровка не представляет особой сложности, однако у представителей трибы Megachilini многочисленные скрытые стерниты прочно соединены друг с другом и латерально с тергитами; они часто тонкие и легко разрываются по средней линии, поэтому успешное извлечение может быть затруднено, и начинающим следует практиковаться на других группах.
У самцов S8 обычно имеет срединную базальную точку или угол прикрепления мышц, отсутствующий на других стернитах; эта структура называется спикулумом (рис. 10-15b). Стерниты и тергиты у обоих полов, за исключением T1 и S1, имеют базолатеральный выступ или аподему с каждой стороны (рис. 10-12).
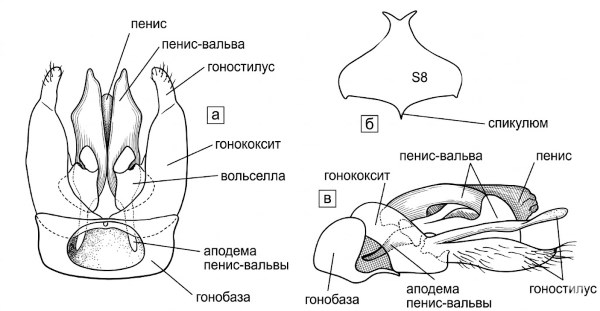
Рисунок 10-15. Генитальные структуры самцов пчел: а — схема гениталий с брюшной стороны; б — схема S8; в — боковой вид гениталий самца Coelioxoides exulans (Holmberg), показывающий верхний и нижний гоностили. а, в модифицированы по Michener, McGinley and Danforth, 1994; в — по Roig-Alsina, 1990.
Мужские гениталии (рис. 10-15а) имеют с каждой стороны, дистальнее основания гонококсита, собственно гонококсит, к дистальному концу которого обычно прикреплен гоностиль. Хотя на гонококсите может быть несколько волосков, гоностиль часто довольно обильно опушен и поэтому легко распознается. В большинстве случаев гонококсит и гоностиль частично срастаются, и их сочленение заметно только с одной стороны области соединения. Часто сращение бывает полным, либо гоностиль утрачивается, так что вместо двусегментного придатка обнаруживается несегментированный выступ. В этом случае бывает невозможно определить, отсутствует ли гоностиль или полностью слился с гонококситом; такая структура называется гонофорцепсом. Обычно дистальная часть гонофорцепса волосистая, и эта часть, вероятно, представляет собой гоностиль. В различных группах можно найти родственные формы обоих типов — одни с отчетливыми гоностилями и гонококситами, другие с полным слиянием или утратой гоностилей.
У многих групп пчел мужской гоностиль разделен до основания, так что с каждой стороны, по-видимому, имеется по два гоностиля, выходящих из смежных частей гонококсита (рис. 10-15с). Они называются верхним (дорсальным) гоностилем и нижним (вентральным) гоностилем. Последний часто отсутствует, и в этом случае верхний гоностиль называется просто гоностилем; именно верхний гоностиль в большинстве литературных источников именуется гоностилем. Нижний гоностиль у разных пчел принимает различные формы и не во всех случаях может быть гомологичен; у таких пчел, как Epicharis и Eufriesea, а также у Coelioxoides (рис. 10-15с) (Apidae), он выглядит как типичный гоностиль — дистально направленный и волосистый.
Обычно основание нижнего гоностиля контактирует с основанием верхнего гоностиля или даже является общим. У некоторых Halictinae нижний гоностиль сходен с верхним, с мелкими волосками, однако у многих других галиктин нижний гоностиль направлен вниз; в таких случаях его обычно называют ретроградной долей. У пчел трибы Allodapini вентроапикальная пластинка, несущая щетинки, похожие на колышки, может быть гомологична нижнему гоностилю.
Волселлы (рис. 10-15а) часто легче всего распознать по сильно склеротизованным темным зубцам — дигитусу и куспису — на противоположных поверхностях. У других перепончатокрылых эти части явно выполняют функцию клешней, но у пчел волселлы редуцированы (часто отсутствуют), и дигитус сливается с телом волселлы и, следовательно, неподвижен. Тем не менее, Снодграсс (Snodgrass, 1941) показал мышцы, которые двигают дигитусом в области парамер и гоностилей. На рис. 10-15а дистальная медиальная волосистая структура — это дигитус, латеральнее которого расположен куспис.
Клапаны пениса (рис. 10-15а) соединены на дорсальной поверхности у их основания перемычкой. У многих Apinae эта перемычка расширяется кзади, образуя дорсальную пластинку, называемую лопаткой. У некоторых пчел часто имеется большой и сложный, в основном перепончатый эндофаллус, который обычно располагается внутри того, что здесь называется пенисом (см. Roig-Alsina, 1993). Род Apis уникален среди всех перепончатокрылых своим огромным эндофаллусом и редукцией всех остальных наружных частей мужских гениталий (рис. 119-3).
Некоторые специальные термины, используемые только для обозначения мужских гениталий Meliponini и определенные в разделе 118, — это амфигональные, ректигональные и шизогональные. Специальные термины, применявшиеся в прошлом для описания генитальных структур Bombini, описаны в разделе 117, особенно в таблице 117-1. Гениталии и скрытые стерниты многих пчел были проиллюстрированы Снодграссом (Snodgrass, 1941) и во множестве таксономических работ, цитируемых в описаниях таксонов пчел в последующих разделах. Особенно большое количество иллюстраций этих структур у различных таксонов можно найти у Сондерса (Saunders, 1882, 1884), Строла (1908), Митчелл (1960, 1962) и Миченера (1954b, 1965b), а различные работы Радошковского также богаты изображениями гениталий.
Представленная здесь детальная морфология скопальных структур, метасомы и гениталий имеет фундаментальное значение для таксономической практики. В последующих разделах систематической части эти признаки будут постоянно использоваться при описании родов и видов, что подчеркивает необходимость единообразного понимания анатомических особенностей. Детальное знание строения собирательного аппарата и терминальных сегментов позволяет не только точно идентифицировать таксоны, но и реконструировать филогенетические отношения, а также понимать адаптации, связанные с образом жизни и репродуктивной биологией различных групп пчел.
Сведения об авторах и источниках:
Авторы: Charles D. Michener
Источник: Пчелы всего мира
Данные публикации будут полезны студентам биологических специальностей, специализирующимся на энтомологии, аспирантам и исследователям в области эволюционной биологии, этологии и генетики насекомых, а также пчеловодам и всем, кто интересуется репродуктивными стратегиями и биологией развития общественных и одиночных пчел.
Дата добавления: 2026-03-15; просмотров: 209;
Поиск по сайту
Узнать еще
- Анатомическая терминология и морфология пчел: строение головы и диагностические признаки
- Артикуляционный аппарат. Изображение голосового аппарата человека
- Архитектура факс-аппарата: микропроцессор, модем и панель управления
- В процессе: Кирпичи и детали, структуры и формы
- Вибрационное опыление и эволюция пчел: механизмы специализации и экологии
- Гаплодиплоидная система определения пола и репродуктивная биология пчел: морфология, развитие и эволюционные стратегии
- Геоморфология и экология приливных отмелей
- Геоморфология речных дельт и ледниковых побережий: формирование, типы и значение

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
