СТРУКТУРА И ФУНКЦИИ ИММУННОЙ СИСТЕМЫ 12 глава
Репертуар распознающих Рц периферических T–лимфоцитов зависит от MHC организма. Исходная биологическая комплементарностъ Рц T–лимфоцитов рассчитана именно на молекулы MHC всех клеток собственного организма.
В функциональном отношении зрелые неиммунные Tab–лимфоциты, выходящие из тимуса, представлены двумя основными субпопуляциями: одна из них несет на мембране молекулу CD8, вторая — CD4. CD8+ T–лимфоциты дифференцированы для выполнения функций цитотоксических T–лимфоцитов(ЦТЛ). ЦТЛ сами непосредственно, своим «клеточным телом» (их называют «перфорин–гранзимовыми киллерами»), убивают клетки (поэтому «ц» — от цито-), на мембране которых они распознали Аг. CD4+ T–лимфоциты дифференцированы в потенциальных продуцентов цитокинов — секретируемых из клетки биологически активных молекул, предназначенных для взаимодействия и вовлечения в процесс других клеток–партнёров по иммунному ответу. CD4+ T–лимфоциты около 30 лет назад назвали лимфоцитами–хелперами (helper — помощник), поскольку эксперименты свидетельствовали, что B–лимфоциты не способны продуцировать АТ без помощи T–лимфоцитов, хотя сами T–лимфоциты не синтезируют иммуноглобулины, но B–лимфоциты без участия T–лимфоцитов их тоже не синтезируют либо совсем, либо синтезируют иммуноглобулины только класса М. За CD4+ T–лимфоцитами закрепилась аббревиатура Th (от T–helper). Однако впоследствии среди Т4–лимфоцитов обнаружены и перфорин–гранзимовые ЦТЛ.
Определённое разнообразие дифференцировки CD4+ T–лимфоцитов в процессе развития иммунного ответа называют иммунным отклонением — отклонением иммунного ответа в сторону той или иной функциональной субпопуляции. В генетически здоровом организме отклонение иммунного ответа зависит от внешних факторов: в первую очередь химических качеств Аг, его дозы, пути попадания во внутреннюю среду, сопутствующих факторов типа адъювантов или инфекций, кратности иммунизации и т.д.
Хотя опухоли — образования патологические, но они позволяют составить представление о нормальных субпопуляциях клеток той или иной ткани. В клинике известны нозологические единицы опухолей из клеток тимуса (табл. 5.3).
Таблица 5.3. Опухоли тимуса: из T–лимфоцитов и АПК
| Болезнь | Тип клеток | Маркёры на мембране | Локализация |
| Острый лимфо-бластный лейкоз | Ранний предшественник лимфоцитов | CD10; CD19; CD20 | Тимус, кровь, костный мозг |
| Тимома | Эпителиальные клетки стромы тимуса | Цитокератины | Тимус |
| Острый лимфо-бластный T–лейкоз | Тимоциты | CD1 | Тимус, кровь |
| Синдром Сезари*; T–лейкоз взрослых (HTL V–1); mycosis fungoides; Хронический T–лейкоз | Та или иная субпопуляция зрелых T–лимфоцитов*** | CD3+/TCR; CD4 или CD8 | Периферические лимфоидные органы, кровь |
| Болезнь Ходжкина** | АПК (моноциты) | CD30 | Периферические ткани |
* Синдром Сезари называют ещё эксфолиативным дерматитом. Он характеризуется сильным зудом, инфильтрацией кожи атипичными мононуклеарными клетками, алопецией, отёком, изменённой пигментацией кожи, изменениями ногтей.
** Болезнь Ходжкина разнообразна по клиническим формам. В основе патогенеза — опухолевая трансформация АПК, вероятнее всего, моноцитов. Но опухолевые АПК стимулируют пролиферацию T–лимфоцитов, и их количество может существенно превосходить нормальные показатели в крови и в тканях. Поэтому болезнь называют лимфомой Ходжкина. В некоторых случаях лимфо-пролиферация отсутствует. Пролиферируют ретикулярные стромальные клетки, и клинически эту форму идентифицируют как узелковый склероз (nodular sclerosis). Прогноз более благоприятен при лимфоме Ходжкина, чем при узелковом склерозе.
***Клональность лимфомы диагностируют анализом генов TCR в опухолевых клетках. Если планируют аутотрансплантацию костного мозга после радио- или химиотерапии, то методом ПЦР важно показать отсутствие генов TCR опухолевого клона в ткани костного мозга.
5.6. Главный комплекс гистосовместимости
«Главный комплекс гистосовместимости» (Major Histocompatibility Complex — MHC) — одно из трудных понятий для начинающих изучать иммунологию. Главная причина этого — оторванность названия от природного предназначения. Эта оторванность объясняется историей развития наших знаний о MHC. Так сложилось, что сначала MHC был выявлен в сугубо искусственной модели трансплантации тканей и органов одного организма во внутреннюю среду другого как некий значительный по размеру комплекс генов, от которых больше, чем от других, зависит скорость отторжения трансплантата. Но поскольку в естественной природе нет процессов трансплантации тканей отдельных особей во внутреннюю среду других особей, много лет имело место молчаливое недоумение — зачем нужны эти гены и продукты, которые они кодируют, организмам, которым не пересаживают и никогда не будут пересаживать чужие органы? Это недоумение разрешили Р. Цинкернагель и П. Дохерти, открывшие в 1973–1974 гг. природные функции белков MHC — представлять для распознавания пептидные Аг для T–лимфоцитов.
Гены MHC были открыты и был введён этот термин в работах 40-х годов в лаборатории, а затем институте Джорджа Снелла — знаменитого иммуногенетика. Иммуногенетики затратили много труда на выведение инбредных линий мышей и подтвердили основные законы трансплантации. Главный закон трансплантации, состоящий в том, что природа не рассчитана на смешение органов уже рожденных особей и пересаженные органы от другого организма в норме всегда отторгаются, люди знали, судя по всему, всегда, сколько существует человеческая культура. Давно знали, что пересадки тканей своего организма в пределах своего организма возможны: пластические операции с аутотрансплантацией описывали средневековые хирурги.
Применительно к генетическим отношениям доноров и реципиентов используют следующие термины:
· аутологичный — свой собственный;
· сингенный (или изогенный) — полностью генетически идентичный организм (трансплантат). Сингенны мыши одного пола одной инбредной линии. Сингенны однояйцевые близнецы;
· аллогенный — другой организм в пределах одного вида. Аллогенны мыши разных инбредных линий;
· конгенный — организм, отличающийся от другого одним (в идеале) геном при одинаковости всех других;
· ксеногенный — организм другого биологического вида.
«Мышиные» иммуногенетики 60-х годов XX в., изучавшие главный комплекс гистосовместимости, несколько иными словами сформулировали те же законы трансплантации, которые в 1912 г. описал Георг Шёне в своей книге «Гетеропластическая и гомопластическая трансплантация»:
Ú аллогенные и ксеногенные трансплантаты всегда отторгаются;
Ú аутологичные и сингенные трансплантаты приживаются.
Гибриды 1–го поколения — F1 (P1´P2) не отторгают трансплантаты обеих родительских линий (P1, P2), но каждая из родительских линий отторгает трансплантат от гибрида (это свидетельствует о кодоминантности в гибридах экспрессии генов, кодирующих продукты, которые являются мишенью отторжения). Кодоминантность означает, что экспрессируются аллели обеих гомологичных хромосом (т.е. от каждого из родителей):
АА´BB ® AB
Ú 50% потомства от обратного скрещивания (F1´P1/2) быстро отторгают трансплантат от второй родительской линии (в случае трансплантации кожного лоскута быстрое отторжение наступает на 8-е сутки). Отсюда по законам менделевской генетики следует, что быстрое отторжение контролирует один локус. Его-то Снелл и назвал главным комплексом гистосовместимости (MHC).
АB´AA ® AA, AA, AA, AB, AB
У мышей локус MHC сегрегируется вместе с геном, кодирующим некий Аг, выявляемый АТ на эритроцитах мышей. Этот Аг в то время называли «Аг–2». Локус, контролирующий гистосовместимость, обозначили Н–2 (Н — от Histocompatibility — гистосовместимость). Верхний буквенный индекс при Н–2 (Н–2а, H–2k, H–2d и т.д.) означает гаплотип данного локуса. У различных инбредных линий гаплотип Н–2 может быть разным или одинаковым.
Локус MHC весьма значителен по размеру — порядка 4´106 пар нуклеотидов (это 2–3 сентиморгана), что соизмеримо с целым геномом бактерии Escherichia coli. Такое количество ДНК у человека содержит около 100 генов. Далеко не все из этих генов и их продуктов описаны и идентифицированы. У мышей выявлены 3 независимых локуса в комплексе MHC — Н–2К, H–2D и H–2L. Позже между ними картировали локусы I–A и I–Е (I от immune response genes — гены иммунного ответа). В 60-х годах Барух Бенацерраф и др. в опытах на морских свинках установили, что интенсивность продукции АТ к белковым Аг контролируется аутосомными генами из комплекса MHC. Аналогичное явление ясно обнаруживалось и в опытах на мышах разных инбредных линий.
Гомологичный комплекс локусов у человека идентифицировали уже другие исследователи (Жан Доссе и др.) и не в экспериментах по трансплантации, а в исследованиях реактивности АТ из крови людей, перенесших трансплантации, или многорожавших женщин с лейкоцитами из крови других людей. Поэтому комплекс обозначили HLA (от Human Leukocyte Antigen — антигены лейкоцитов человека). Первые 3 Аг, выявленные в этих серологических реакциях (и соответствующие гены), обозначили HLA–A, HLA–B, HLA–C. Следующий локус обнаружили не серологически, а по индукции пролиферации в смешанной культуре лимфоцитов донора и реципиента; его обозначили HLA–D. Следующий идентифицированный серологически локус был близко сцеплен с HLA–D, его обозначили HLA–DR (от D–related). Следующие два локуса обозначили соседними с R буквами P и Q — HLA–DP и HLA–DQ.
Позже, когда выяснили, какие именно белковые продукты кодируют разные структурные гены из комплекса MHC, комплекс разбили на два класса — I и II — MHC–I, MHC–II. К классу I у человека отнесли локусы HLA–A, HLA–B и HLA–C, к классу II — соответственно HLA–DR, HLA–DP и HLA–DQ. Схема расположения в геноме локусов комплекса MHC человека показана на рис. 5.5.

Рис. 5.6. Расположение генов MHC человека в хромосоме 6 (схема).
Продукты генов локусов MHC–I — мембранные белки, конститутивно экспрессированные на всех ядросодержащих клетках тела, кроме сперматозоидов, но в разных количествах на клетках различных гистотипов: больше всего этих молекул на лимфоцитах и лейкоцитах. MHC–II — тоже мембранные молекулы, но экспрессированы не на всех, а только на некоторых гистотипах клеток — на дендритных клетках, B–лимфоцитах, моноцитах/макрофагах, эндотелии сосудов. Первые 3 типа клеток относят к так называемым профессиональным АПК — это дендритные клетки, B–лимфоциты и макрофаги, поскольку на этих клетках, кроме молекул MHC–II и I, экспрессируются все необходимые корецепторные молекулы и цитокины, достаточные для активации T–лимфоцита к иммунному ответу (табл. 5.4).
Таблица 5.4. Относительные уровни экспрессии MHC–I и II на разных клетках
| Клетки | MHC–I | MHC–II |
| T–лимфоциты | +++ | + (у человека) |
| B–лимфоциты | +++ | +++ |
| Дендритные клетки | +++ | +++ |
| Макрофаги | +++ | + |
| Эпителий тимуса | + | +++ |
| Эндотелий | + | + |
| Нейтрофилы | +++ | Нет |
| Гепатоциты | + | Нет |
| Клетки почки | + | Нет |
| Нейроны мозга | + | Нет |
| Эритроциты | Нет | Нет |
Примечание. Количественная выраженность того или иного свойства представлена в условных знаках: «+» — немного; «++» — больше, чем «+"; «+++» — больше, чем «++».
Эндотелий тоже выполняет функции АПК, но особого предназначения. Вероятно (есть такие экспериментальные данные), экспрессия комплексов Аг–пептидов с MHC на клетках эндотелия является тем специфическим сигналом, который «заманивает» иммунные лимфоциты из циркулирующей крови в больное, поражённое патогеном место в тканях (обеспечивает антигенспецифичный хоминг).
Каждый локус MHC–I кодирует полипептидную цепь, обозначаемую традиционно a, с мол. массой около 44 тыс., примерно из 325 остатков АК. Цепь a имеет 3 внеклеточных домена (a1, a2, a3), трансмембранный участок и цитоплазматический участок из 55 АК–остатков. С внеклеточными a–доменами нековалентно ассоциирована лёгкая полипептидная цепь с мол. массой 12 тыс., называемая b2–микроглобулином. Микроглобулин b2 кодируется геном, не сцепленным с MHC. Домены a1 и a2 формируют углубление (щель) размером около 2,5 нм. Именно в этом углублении располагается пептид–Аг, предназначенный для распознавания T–лимфоцитами. Пептид ни в коем случае не сорбируется на MHC извне клетки, а конъюгирует с a–цепью нековалентными связями, но с достаточной аффинностью (Kd »10–6 М) внутри клетки при фолдинге (сворачивании, т.е. изменении конформации MHC). Как и все трансмембранные белки, MHC–I гликозилированы; места гликозилирования a–цепи — Asn 86, Asn 176, Asn 256.
MHC–I связывает пептиды длиной 8–10 АК–остатков, фиксируя пептид по обоим концам молекулы — С и N. Молекулы MHC разных аллельных вариантов связывают пептиды с определёнными остатками АК в так называемых якорныхпозициях: это C–концевой остаток и 2-й или 5-й остаток с N–конца. Важно, что каждый конкретный аллельный вариант молекулы MHC связывает пептиды с данными, а не другими якорными остатками АК. Другой аллельный вариант MHC имеет химическое сродство к иным якорным остаткам АК. Поэтому важно посчитать, сколько же вариантов MHC имеет каждый отдельный человек, так как только на те пептиды, которые способны связать MHC особи, данная особь потенциально способна развить защитный иммунный ответ. Если же некий Аг не связывается ни с одним вариантом MHC из тех, которыми располагает особь, то против этого Аг не будет иммунного ответа, организм беззащитен перед таким Аг. Отсюда понятно, что генетический контроль иммунного ответа сильно зависит не только от генов, специально экспрессирующихся в лимфоцитах и АПК, но и от генов, экспрессирующихся во всех клетках организма, — MHC. Исключение составляют безъядерные зрелые эритроциты и сперматозоиды:на них нет ни MHC–I,ни MHC–II.
Белки MHC–II (класса II) — мембранные молекулы — гетеродимеры из двух трансмембранных гликопротеинов: a–цепь (мол. масса 34 тыс.) и b–цепь (мол. масса 29 тыс.). Внеклеточная часть каждой цепи имеет два домена. Пептидсвязывающее углубление (щель) формируют совместно a1- и b1–домены, в отличие от аналогичного углубления в составе MHC–I щель MHC–II открыта с обеих сторон (рис. 5.7). Стереохимическая открытость углубления MHC–II позволяет связывать более длинные пептиды, чем в случае MHC–I, — до 30 остатков АК. Однако чаще всего из комплексов с MHC–II удаётся диссоциировать пептиды размером 13–17 остатков АК. Якорные остатки для большинства изученных аллельных вариантов MHC–II находятся в позициях 1; 4; 6 и 9.
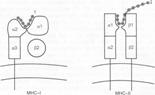
Рис. 5.7. Структура молекул MHC–I и MHC–II (схема). Молекулы MHC–I — трансмембранные гетеродимеры: a–цепь большая, имеет три внеклеточных домена. «Щель» для связывания пептидом формируют два терминальных домена a–цепи (a1 и a2). b–Цепь малого размера, в мембрану не внедряется, с a–цепью ассоциирована нековалентно и её называют b2–миκроиммуноглобулином. Молекулы MHC–II также трансмембранные гетеродимеры. Полипептидные цепи a и b равновелики, обе пенетрируют мембрану, у обеих по два внеклеточных домена и «щель» для связывания пептидов формируют внешние домены обеих цепей. 1 — пептид–Аг в комплексе с MHC–I; 2 — пептид–Аг в комплексе с MHC–II.
5.7. Механизмы образования комплексов «пептид–антиген» с молекулами главного комплекса гистосовместимости
В любой эукариотической клетке есть две «зоны», или, как их принято называть от английского, компартменты. Одна зона связана с внешней внеклеточной средой и это зона мембранных структур и везикул, включающая в себя аппарат Гольджи, эндоплазматический ретикулум, лизосомы, эндосомы и фагосомы. Пептиды, образующиеся в данной зоне, обслуживают синтезируемые клеткой молекулы MHC–II. Происхождение пептидов везикул внеклеточное: они образуются в результате протеолиза белков, захваченных клеткой посредством эндоцитоза или фагоцитоза. Эндосомы или фаголизосомы, сливаются с мембранными структурами, на которых фиксируются вновь синтезированные на полирибосомах молекулы a- и b–цепей MHC–II и ещё одна так называемая инвариантная полипептидная цепь — Ii. Эта цепь «прикрывает» MHC–II от связывания с пептидами внутри эндоплазматического ретикулума и в дальнейшем обеспечивает экспозицию молекул MHC–II внутрь везикул — эндосом или фаголизосом. Комплекс из 3 тримеров abIi в эндоплазматическом ретикулуме удерживается связью с кальнексином. В мембранных внутриклеточных структурах есть специальный компартмент М–II–С, в который физически упираются эндосомы и лизосомы с поглощённым внеклеточным содержимым. И только с участием пептида молекула MHC–II принимает правильную конформацию, продвигается к мембране и экспрессируется на ней. «Чистые» молекулы MHC классов I и II на мембране клетки не экспрессируются. Если клетка не располагает достаточным количеством чужого пептидного материала, то молекулы MHC–II связывают короткий фрагмент, образующийся при протеолитическом расщеплении инвариантной цепи — CLIP. В наслоении пептида на MHC–II участвует ещё одна молекула — HLA–DM. HLA–DM на какое-то время стабилизирует пустые молекулы MHC–II (без пептида). Отсюда понятно, почему представление Аг именно молекулами MHC–II происходит при развитии защитных иммунных реакций на внеклеточные и везикулярные инфекции. Поскольку комплексы антигенов–пептидов с молекулами MHC–II распознают исключительно CD4+ T–лимфоциты, то и в защите от внеклеточных и везикулярных инфекций главную роль играют реакции, обеспечиваемые именно CD4 T–лимфоцитами (рис. 5.8).
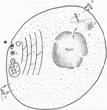
Рис. 5.8. Строение клетки и «зоны обслуживания» молекулами MHC–I и MHC–II (схема). 1 — зона (или компартмент) мембранных структур клетки (везикулы, эндоплазматический ретикулум, аппарат Гольджи). Эта зона непосредственно сообщается с внеклеточной средой; 2 — зона (или компартмент) цитозоля непосредственно сообщается с внутриядерным содержимым; 3 — протеасомы в цитозоле; 4 — фагосомы; 5 — фаголизосомы сливаются с везикулами, содержащими несконформированные молекулы MHC–II; 6 — комплекс MHC–II — пептид «внеклеточного» происхождения экспрессирован на мембране клетки; 7 — несконформированные молекулы MHC–I в зоне досягаемости пептидов с протеасом; 8 — комплекс MHC–I — пептид «внутриклеточного происхождения» экспрессирован на мембране клетки.
Вторая «зона» в клетке — зона цитозоля — неструктурированного мембранами внутриклеточного содержимого. Цитозоль непосредственно сообщается через крупные ядерные поры с содержимым ядра. В ядре идут процессы транскрипции РНК с ДНК (как нормальные клеточные, так и вирусные при вирусных инфекциях клеток). РНК из ядра транспортируется для трансляции белка в цитозоль на полирибосомы, и для фолдинга (принятия правильной конформации) белков под контролем шаперонов («катализаторов» принятия полипептидной цепью биологически значимой конформации) полипептидные цепи направляются в специальные участки эндоплазматического ретикулума. Лишние, испорченные, неправильно конформированные белки постоянно расщепляются в цитозоле так называемыми протеасомами. Протеасомы — это мультипротеазные комплексы из 28 СЕ, каждая из которых имеет молекулярную массу 20 тыс. или 30 тыс. Два из 3 вариантов СЕ протеасом — LMP2 и LMP7 — кодируются генами, расположенными внутри комплекса MHC.
Протеасомы осуществляют зависимую от убиквитина деградацию белков цитозоля. И именно пептиды, образующиеся в результате работы протеасом, поступают в «поджидающие» их в нужном месте эндоплазматического ретикулума ещё несконформированные молекулы MHC–I: вступая в связь с пептидом, молекулы MHC–I принимают биологически правильную конформацию и направляются для экспрессии на клеточную мембрану. Исследование мутаций и генетических дефектов, приводящих к иммунодефицитным состояниям, выявило зависимость представления Аг молекулами MHC–I от двух АТФ–связывающих транспортёров TAP–1 и TAP–2. В мембране эндоплазматического ретикулума эти два полипептида формируют гетеродимер, ориентированный гидрофобными участками в полость ретикулума, а АТФ–связывающими доменами — в сторону цитозоля. Именно они доставляют пептиды для связывания с молекулами MHC–I. TAP имеют сродство к пептидам с гидрофобными или основными остатками АК на C–конце. Гены TAP–1 и TAP–2 расположены внутри комплекса MHC.
Вновь синтезированная a–цепь MHC–I вступает в связь с мембранным протеином эндоплазматического ретикулума кальнексином (мол. масса 88 тыс.), который обеспечивает частичный фолдинг a–цепи. Кстати, тот же кальнексин образует комплексы с вновь синтезированными и частично сконформированными полипептидами MHC–II, TCR, иммуноглобулины (т.е. всеми иммунологически значимыми молекулами). При присоединении к a–цепи b2–микроглобулина связь с кальнексином диссоциирует, и комплекс ab2 присоединяется для дальнейшего прохождения процесса приобретения конформации к кальретикулину эндоплазматического ретикулума и TAP–1–ассоциированному протеину тапасину. Таким образом создаются условия для соединения пептида–Аг с принимающим конформацию комплексом MHC–I. При генетических дефектах в TAP молекулы MHC деградируют внутри клетки, не попадая на клеточную мембрану. В норме в клетке молекулы MHC–I присутствуют в избытке по отношению к пептидам. Поэтому в клетке при вирусных инфекциях довольно быстро пептиды вирусного происхождения оказываются представленными на поверхности заражённых клеток в составе комплексов с MHC–I.
Молекулы MHC–I представляют для распознавания T–лимфоцитам внутриклеточно синтезированные пептидные Аг. В плане защиты от инфекции этот механизм работает применительно в первую очередь к вирусным, а также цитозольным бактериальным внутриклеточным инфекциям. Поэтому CD8+ T–лимфоциты «рассчитаны» в первую очередь на обеспечение противовирусной защиты, так как в комплексе с MHC–I именно и только CD8 T–лимфоциты распознают пептидные Аг.
В отсутствие инфекций молекулы MHC–I и MHC–II формируют комплексы с эндогенными (собственными) пептидами. Обратим внимание на то, что дифференцировка T–лимфоцитов в тимусе в норме в стерильном изнутри организме идёт при взаимодействии отбираемых TCR со своими пептидами, а не на отборе «своего и чужого». Поэтому естественно, что среди случайно формируемого разнообразия вариантов антигенсвязывающих структур даже после позитивной и негативной селекции тимоцитов среди выходящих на периферию T–лимфоцитов обязательно найдутся TCR, способные специфично связываться, наверное, со всеми собственными «Аг». Опыт показывает, что так оно и есть. Но деструктивный иммунный ответ на собственные Аг в норме не развивается, и в дальнейшем мы обсудим то, что известно о механизмах периферической толерантности.
Стабильное связывание Аг молекулами MHC внутри клетки при формировании собственной конформации, а не простая сорбция из раствора Аг снаружи уже экспрессированными на мембране молекулами MHC, обеспечивает направление T–лимфоцитарного иммунитета на реально инфицированные клетки тела.
Иммунное воспаление, управляемое активными T–лимфоцитами, — это наиболее «энергичное» воспаление по сравнению с другими вариантами, и такое воспаление нельзя «натравливать» на ложные мишени, иначе организм будет не направленно избавляться от инфекции, а страдать от неоправданно разлитой альтерации тканей.
Взаимодействие TCR с комплексом «пептид–MHC–I/II» изображено на рис. 5.9.
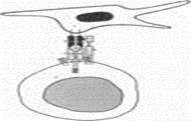
Рис. 5.9. Взаимодействие TCR с комплексом «MHC–пептид» на антигенпредставляющей клетке (АПК) 1 — АПК (например, дендритная); 2 — CD4+ T–лимфоцит; 3 — MHC–II; 4 — пептид (собственно Аг); 5 — a- и b–цепи TCR; 6 — молекула CD4.
5.8. Суперантигены
В борьбе за своё существование инфекционные микроорганизмы «научились» синтезировать некие вещества, которые поступают с «нашими» T–лимфоцитами «не по правилам». Эти вещества называют суперантигенами. Когда они попадают в организм, то последний не может встретить их попыткой распознавания специфическим TCR потому, что суперантигены благодаря своим особым химическим свойствам способны присоединяться снаружи клетки (т.е. без процессинга в АПК) с одной стороны к молекулам MHC–II, но не в области «клефта для пептида», а «сбоку», как правило, к a–цепи, и одновременно к V–области b–цепи TCR на T–лимфоците.
Это связывание вызывает активацию T–лимфоцитов, но не антигенспецифичного клона, а поликлональную. Обычно суперантигены активируют 2–20% периферических CD4+ T–лимфоцитов. В результате многие T–лимфоциты продуцируют и секретируют активационные цитокины, что вызывает синдром общей интоксикации в организме. А затем активированные T–лимфоциты погибают путём апоптоза. Таков закон лимфоцитарного иммунитета: на свой ли Аг или неспецифически, но единожды активированный лимфоцит обречен на апоптоз (феномен индуцированной активацией клеточной смерти). (Феномен иммунологической памяти — особое явление, и хотя оно исключительно характерно именно для лимфоцитарного иммунитета, механизмы его непонятны до настоящего времени). Но фактом является то, что после поликлональной активации лимфоцитов наступает физический дефицит лимфоцитов, следовательно, иммунодефицит.
Известны суперантигены бактериального и вирусного происхождения, такие как энтеротоксины стафилококков, токсин синдрома токсического шока, мембранный белок вируса опухолей молочных желёз мышей, мало охарактеризованные суперантигены вирусов Эпштейна–Барр, вируса бешенства, вирусов иммунодефицита человека (ВИЧ) и животных и многие другие. Клиника пищевых отравлений, например, обязана главным образом действию энтеротоксинов стафилококков на T–лимфоциты. Иммунодефицит в ранние сроки (и в поздние тоже) ВИЧ–инфекции в значительной мере объясняется лимфотоксическим действием суперантигенов ВИЧ.
5.9. Генетический полиморфизм главного комплекса гистосовместимости
Итак, в случае T–лимфоцитов комплекс «пептид–Аг» связывается не только с Рц T–лимфоцита для Аг, но и (причём по времени «до» T–лимфоцита) с молекулами MHC Аг–представляющих клеток. Разнообразие TCR природа создаёт комбинаторикой сегментов ДНК — уникальным процессом рекомбинации ДНК в соматических клетках. А как молекулы MHC приспособлены к открытому множеству заранее неизвестных Аг? Иначе, чем TCR и иммуноглобулины. Рекомбинации ДНК в генах MHC нет. Каждый человек наследует определённый набор генов MHC, один и тот же на всю жизнь. Гены MHC кодоминантны. Это значит, что на равных экспрессируются гены обеих гомологичных хромосом — от матери и от отца.
Гены MHC полигенны, т.е. существует не один, а несколько локусов классов I и II внутри комплекса MHC. Ещё есть и так называемые неклассические гены, правильнее их было бы назвать не принадлежащими к ранее охарактеризованным классам MHC I и II. Сколько же генов MHC? Этот участок генома картирован не полностью, но то, что известно о нем, показано на рис. 5.5.
Таким образом, генов MHC–I по 3 на каждой из гомологичных хромосом (А, В, С), следовательно, в сумме их 6. Генов MHC–II также по 3 на каждой из гомологичных хромосом (DP, DQ, DR), следовательно, в сумме их тоже 6. Если у матери и отца нет одинаковых аллелей, то каждый человек имеет 12 генов MHC классов I и II, вместе взятых. Неклассических генов MHC в области расположения генов класса I описано 6 (Е, F, G, Н, J, X). Возможно, будут открыты и новые гены. Неклассические гены MHC, видимо, неполиморфны или не столь полиморфны, как гены классов I и II.
Известны гены за пределами MHC, кодирующие молекулы, которые обеспечивают представление Аг T–лимфоцитам. Примером такой молекулы (и гена соответственно) является CD1, способная связывать и представлять T–лимфоцитам небелковые Аг (например, миколевую кислоту и липо-арабиноманнан оболочки микобактерий).
Дата добавления: 2016-07-18; просмотров: 2154;
Поиск по сайту
Узнать еще
- a-спираль b-складчатая структура
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
