СТРУКТУРА И ФУНКЦИИ ИММУННОЙ СИСТЕМЫ 10 глава
Ответы клеток–мишеней происходят при сочетании двух феноменов — пролиферации и биосинтеза специфических белков.
О том, что проведение сигнала внутрь клетки — не только конечная цель, но и необходимо для самого процесса дифференцировки, свидетельствует тот факт, что генетический дефект в тирозинкиназе Btk имеет следствием иммунодефицитную патологию с полным отсутствием у человека B–лимфоцитов — Х–сцепленную агаммаглобулинемию Брутона.
4.8. Стадии лимфопоэза B–лимфоцитов
Введение в зародышевые клетки искусственного перестроенного трансгена тяжёлой цепи полностью подавляет перестройку одноимённых собственных генов клетки. Но если трансген не содержит кода для трансмембранного участка тяжёлой цепи, то трансген не мешает перестройке собственного одноимённого гена. Следовательно, для того чтобы в конечном счёте дифференцированный B–лимфоцит имел строго один вариант тяжёлой цепи и один вариант лёгкой, ещё в процессе дифференцировки необходима экспрессия тяжёлой цепи на мембране. Так оно и есть. Как только в клетке произошла трансляция полипептида тяжёлой цепи, он экспрессируется на мембране в составе так называемого пре–B–Рц. Чтобы это могло случиться, в про–B–клетке синтезируются два специальных полипептида, которые нековалентно соединяются друг с другом, образуя суррогатную лёгкую цепь. Один из этих полипептидов — d5, второй — VpreB. Таким образом, пре–B–клеточный Рц состоит из d5 + VpreB + m–цепь + Iga + Igb. Его экспрессия транзиторна, но абсолютно необходима для правильной дифференцировки B–лимфоцитов. После экспрессии пре–B–Рц временно инактивируются белки RAG, и клетки вступают в процесс интенсивной пролиферации, которая прекращается с исчезновением этого Рц. После завершения этой волны пролиферации вновь экспрессируются гены RAG1 и RAG2 и начинается перестройка генов лёгкой цепи. Как только это произойдет, на развивающемся B–лимфоците будет экспрессироваться дефинитивный BCR состава: L–цепь + m–цепь + Iga + Igb. Эту стадию развития называют незрелым B–лимфоцитом.
Маркёром завершения B–лимфопоэза — образования зрелого неиммунного B–лимфоцита, готового к выходу из костного мозга в периферическую лимфоидную ткань, — является одновременная экспрессия (коэкспрессия) на мембране 2 типов BCR — с IgM и IgD (задействуется альтернативный сплайсинг РНК–транскрипта с md–гена тяжёлой цепи), причём IgD больше, чем IgM.
Прежде чем произойдет экспрессия на мембране IgD, но после того как произошла экспрессия BCR с полноценным IgM, в развитии B–лимфоцитов «предусмотрен» существенный и обязательный этап дифференцировки — селекция (апоптоз) аутореактивных клоновв местах прохождения лимфопоэза, т.е. на территории костного мозга. В природе устроено так, что связывание Аг незрелой B–клеткой, на которой есть антигенраспознающий Рц с IgM, но ещё нет Рц с IgD, является сигналом для апоптоза, т.е. запрограммированной гибели клетки. Таким образом, из случайного репертуара по Аг–связывающим Рц на исходе лимфопоэза убираются B–лимфоциты, несущие Рц, способные с высокой аффинностью связывать белки собственных клеток и растворимые белки, присутствующие в достаточных количествах на территории костного мозга. Такой механизм толерантности к своему называют делецией клона.
Толерантностьюв иммунологии называют отсутствие иммунного ответа конкретной особи на тот или иной (те или иные) Аг, на который(ые) другие особи либо та же особь, но при иных конкретных условиях онтогенеза, потенциально способны развивать иммунный ответ. Делеция клона — не единственный механизм установления толерантности к конкретному Аг со стороны B–лимфоцитов. Известно ещё два механизма: развитие состояния ареактивности (или анергии) и «редакция» Рц по антигенной специфичности. Эти два механизма действуют в периферических лимфоидных тканях.
Реальность механизма делеции клона, специфичного к мультивалентному Аг, который экспрессирован на мембранах собственных клеток, показана в экспериментах с трансгенными мышами. Например, в таком эксперименте, в котором трансгены — гены лёгкой и тяжёлой цепи молекулы иммуноглобулина, специфичного к молекулам Н–2Кb MHC–I. У таких мышей благодаря описанным выше закономерностям все B–лимфоциты имеют один и тот же BCR с иммуноглобулином, кодируемым трансгеном. Если мыши–реципиенты такого трансгена в собственном организме не имеют Аг–мишени (т.е. Н–2Кb), у них нормальное количество B–лимфоцитов в периферических лимфоидных тканях, только все они с одним и тем же Рц. Но если мыши-реципиенты сами имеют ген/Аг Н–2Кb, то у них находят нормальное количество пре–B–клеток, которые, однако, все погибают на территории костного мозга апоптозом и в периферических лимфоидных тканях B–лимфоцитов совсем нет. По тому же механизму клональной делеции погибают и B–лимфоциты на периферии, если они несут Рц, способный связывать молекулы мембран клеток, которые представлены в большом количестве в тех или иных тканях (например, в печени).
Если незрелый B–лимфоцит(с IgM, но без IgD) связывает растворимый Аг (например, в организме дважды трансгенных мышей: один трансген кодирует синтез растворимого белка, второй — АТ к нему), то лимфоцит не элиминируется апоптозом, а остаётся в организме, но приобретает состояние анергии: в результате связывания Аг с Рц не наступает активации лимфоцита к иммунному ответу, наоборот, развивается блок проведения сигнала.
У зрелых B–лимфоцитовв периферических лимфоидных тканях на большие дозы растворимых Аг и особенно при отсутствии адекватного взаимодействия с T–хелперами так же развивается состояние анергии. Такие клетки долго не живут и в течение нескольких дней все равно погибают.
Для полноценной реакции на Аг лимфоциту мало только Рц для Аг. У лимфоцитов есть ещё такой обязательный фактор, как корецепторный комплекс мембранных молекул, связанных с внутриклеточными системами проведения сигналов. Не на каждом Аг есть повторяющиеся эпитопы, следовательно, не каждый Аг способен вызвать перекрестную сшивку (или агрегацию) BCR. Вот в этих случаях и нужны дополнительные сигналы. В корецепторный комплекс B–лимфоцитов входят по крайней мере такие мембранные молекулы, как CD19/CR2 (CD21)/TAPA–1.
Молекула CD19 экспрессирована на всех B–клетках, начиная с ранних стадий лимфопоэза. Генетический нокаут мышей пo CD19 приводит к выраженному дефициту B–клеточного иммунного ответа на любой Аг. Точные механизмы участия CD19 в активации B–лимфоцита Аг неизвестны. В мембране CD19 физически ассоциирована с Рц 2–го типа для компонентов комплемента — CR2 (CD21). Связывание CR2 с компонентами комплемента имеет следствием фосфорилирование молекулы CD19 ассоциированными с BCR киназами. Фосфорилированная молекула CD19 связывает фосфатидилинозитол–3–киназу и молекулу vav (многофункциональная молекула проведения внутриклеточных сигналов), которые усиливают активационные реакции, инициированные с BCR. Физически в мембране к CD19 и CR2 примыкает TAPA–1 (CD81), но роль этой молекулы неизвестна.
В результате только совместной работы BCR и корецепторного комплекса возможен запуск в клетке таких процессов, как пролиферация, экспрессия на мембране молекул, необходимых для взаимодействия с T–лимфоцитами, экспрессия на B–лимфоцитах молекул MHC–II, необходимых для представления B–лимфоцитом Аг T–лимфоциту для распознавания.
Приведём краткие суммированные сведения о динамике приобретения признаков дифференцировки во время развития B–лимфоцитов (лимфопоэза) (табл. 4.3).
Таблица 4.3. Стадии (признаки) дифференцировки B–лимфоцитов в лимфопоэзе
| Стадия развития B–лимфоцита | Состояние генов тяжёлой цепи | Состояние генов лёгкой цепи | Экспрессия белков | Мембранные маркёры |
| Общая лимфоидная клетка–предшественница | Зародышевая конфигурация | Зародышевая конфигурация | RAG1/RAG2 | CD34; CD45 |
| Ранняя про–B–клетка | Перестройка DJ | То же | RAG1/RAG2; TdT; VpreB; | CD34; CD45; CD10; CD19; CD38; MHC–II |
| Поздняя про–B–клетка | Перестройка V–DJ | То же | TdT; VpreB; | CD45R; CD10; CD19; CD38; CD20; CD40; MHC–II |
| Большая пре–B–клетка | Перестройка V–DJ | Перестроенный VDJ | RAG1/RAG2; VpreB; | CD45R; MHC–II; Pre–BCR; CD19; CD20; CD38; CD40 |
| Малая пре–B–клетка | То же | Перестройка V–J | Тяжёлая | CD45R; MHC–II; CD19; CD20; CD38; CD40 |
| Незрелая B–клетка | Перестроенный VDJ в | Перестроенный VJ | CD45R; MHC–II; CD19; CD20; CD40; BCR–IgM | |
| Зрелая неиммунная B–клетка | Экспрессия IgM и IgD | Экспрессия лёгкой цепи | CD45R; MHC–II; CD19; CD20; CD21; CD40; BCR–IgM/BCR–IgD |
После распознавания Аг и вступления в иммунный ответ в периферических лимфоидных органах и тканях B–лимфоцит пройдет ещё две стадии додифференцировки, которые называют иммуногенезом. На второй из этих стадий произойдет дихотомия: B–лимфоцит станет либо B–лимфоцитом памяти (уйдет в дифференцированный резерв на случай, если тот же Аг попадёт во внутреннюю среду организма повторно), либо плазматической клеткой — терминальным продуцентом больших количеств секретируемого иммуноглобулина заданной специфичности. Собственно иммуногенез будет описан ниже, здесь, забегая вперед, приведём признаки дифференцировки B–лимфоцитов на разных стадиях иммуногенеза в B–лимфоциты памяти и плазматические клетки (табл. 4.4).
Таблица 4.4. Признаки дифференцировки B–лимфоцитов в процессе развития иммунного ответа (иммуногенеза)
| Стадия развития B–лимфоцита в иммуногенезе | Состояние генов/РНК/белка тяжёлой цепи | Состояние генов/РНК/ белка лёгкой цепи | Экспрессия особых белков | Маркёры клеточной мембраны |
| Лимфобласт | Альтернативный сплайсинг РНК, биосинтез секретируемой формы | Продуктивный синтез лёгкой цепи | Специфическое АТ | CD45R; MHC–II; CD19; CD20; CD21; CD40 |
| B–лимфоцит памяти | Произошло переключение класса тяжёлой цепи с | В V–генах произошли гипермутирование и отбор наиболее аффинных АТ | – | CD45?; MHC–II; BCR–IgG или IgA, или IgE; CD19; CD20; CD21; CD40 |
| Плазмоцит | Массовая продукция тяжёлых цепей заданного типа | Массовая продукция лёгких цепей заданного типа | Массовая продукция цельных иммуноглобулинов в секреторной форме | Аг плазмоцитов–1 (CD38) |
Третий механизм избегания аутореактивности B–лимфоцитов, «редакция» Рц, используется, по-видимому, в небольшом проценте незрелых B–клеток, в которых ещё активны гены перестройки RAG1 и RAG2. В этих клетках связывание IgM в составе BCR на поверхности незрелого B–лимфоцита с Аг является сигналом для запуска нового процесса рекомбинации VDJ/VJ на второй из двух гомологичных хромосом: если «повезет», то для второго варианта VDJ/VJ на территории костного мозга не найдется Аг и B–лимфоцит выживет и будет иметь шанс быть использованным на периферии.
В костном мозге молодых здоровых мышей ежедневно вступают в митоз 30–40´106 клеток. Из них только 10–15´106 клеток (меньше половины) выходит на периферию. Столько же периферических B–лимфоцитов ежедневно отмирает. Это ежедневное обновление пула составляет 5–10% общего числа периферических B–лимфоцитов. Если «новый» B–лимфоцит по каким-то причинам не попал в лимфоидный фолликул периферических лимфоидных тканей, то его время полужизни не превысит 3 дней. В лимфоидных фолликулах неиммунные B–лимфоциты имеют время полужизни от 3 до 8 нед: все это время они готовы встретить «свой» Аг и вступить в процесс иммуногенеза, т.е. развития иммунного ответа, и стать либо плазмоцитом, либо B–лимфоцитом памяти.
Специализированное анатомическое место пребывания B–лимфоцитов в периферической лимфоидной ткани — фолликулы. В фолликулах B–лимфоциты удерживаются связями со специальными клетками стромы — дендритными клетками фолликулов. ФДК — это не те же самые дендритные клетки, которые присутствуют в покровных тканях [под названием клеток Лангерганса (белых отростчатых эпидермоцитов)], в тимусе (интердигитирующие) и циркулируют в крови. Это другие по гистогенетическому происхождению клетки, скорее всего, ФДК не имеют костномозговое происхождение.
На ФДК экспрессированы Рц для иммуноглобулинов — Fc–Рц (FcR), отличающиеся двумя особенными свойствами. Первое, самое необычное, заключается в том, что, связав однажды комплекс Аг–АТ через FcR, ФДК способны нести его на себе продолжительное время (дни, месяцы, возможно, годы). Второе свойство FcR ФДК — комплекс Аг–АТ не поглощается внутрь клетки.
Именно в фолликулах в связи с ФДК B–лимфоцит, распознавший свой Аг и вступивший ещё при прохождении через T–зависимую парафолликулярную зону лимфатического узла (или другого периферического лимфоидного органа) в адекватное взаимодействие с также распознавшим Аг T–лимфоцитом, интенсивно пролиферирует. На этой стадии развития B–лимфоциты называют центробластами.
В центробластах происходит уникальное даже среди лимфоцитов явление — возрастание аффинности АТ в отношении своего Аг. В дифференцировке T–лимфоцитов аналогичного процесса нет.
Феномен возрастания аффинности АТ по мере прогрессивного развития иммунного ответа — пример классического дарвиновского процесса на клеточном уровне. Уникальной особенностью молекулярной генетики B–лимфоцитов является запрограммированность на повышенную частоту соматических мутаций в уже перестроенных V–генах иммуноглобулинов: в центробластах происходит замена одной пары нуклеотидов (п.н.) на каждые 103 п.н. на 1 митоз. Во всяком другом участке ДНК такое событие случается с вероятностью 10–12, т.е. на 9 порядков реже. Получается, что каждый второй центробласт несет мутацию в V–области молекулы иммуноглобулина. Чем выше оказывается сила связи иммуноглобулина в составе BCR (больше аффинность) с Аг, присутствующим в фолликуле на поверхности ФДК, тем больше вероятность выживания данного B–лимфоцита, ибо на этом этапе дифференцировки связь с Аг является антиапоптозным сигналом на выживание — происходит индукция экспрессии антиапоптозного гена bcl–2. Если в результате генетической или эпигенетической аномалии имеет место повышенная экспрессия антиапоптозных генов, то развивается процесс, который называют лимфопролиферативным, т.е. возникают опухоли из центробластов.
В терминальной стадии дифференцировки B–лимфоцита, в плазмоците, сильно развит эндоплазматический ретикулум. Более 20% всего белкового синтеза плазмоцита составляют секретируемые иммуноглобулины. На мембране плазмоцита иммуноглобулинов уже нет. На плазмоцитах нет и MHC–II; в них невозможно уже переключение классов иммуноглобулинов, невозможно соматическое гипермутирование иммуноглобулиновых генов. Продукция АТ плазматической клеткой уже не зависит от контакта с Аг, не зависит и от взаимодействий с T–лимфоцитами. Первые плазматические клетки локализуются на территории лимфатического узла, в котором инициирован иммунный ответ, а именно в мозговых тяжах; эти клетки вырабатывают АТ для «внутреннего пользования»: данные АТ связывают Аг в комплекс и фиксируются на FcR фолликулярных дендритных клеток. По мере прогрессивного иммунного ответа прошедшие аффинное созревание B–лимфоциты превращаются в плазмоциты, которые мигрируют из лимфатического узла в костный мозг или в собственную пластинку слизистой оболочки, где живут и работают в течение почти 4 нед. Этим сроком и ограничена продолжительность продуктивного гуморального иммунного ответа.
Для всех описанных стадий развития B–лимфоцитов описаны и соответствующие опухоли, сохраняющие и локализацию, и фенотипические признаки исходной нормальной клетки. Опухоли из лимфоцитов всегда клональны, судя по перестроенным генам иммуноглобулинов, т.е. происходят из одной клетки. Именно на опухолях из B–лимфоцитов была изучена молекулярная генетика иммуноглобулинов. Плазмоцитомы (син. миеломы) локализуются в костном мозге. Хотя они являются опухолями из зрелых клеток, но растут, как правило, агрессивно, так как микроокружение изобилует факторами роста.
В типичных случаях нозологический диагноз лейкоза соответствует той или иной нормальной субпопуляции или стадии развития B–лимфоцитов (табл. 4.5).
В опухолевых лимфоидных клетках часто обнаруживают транслокации, при которых локусы генов иммуноглобулинов оказываются физически приближёнными к генам регуляции пролиферации.
Таблица 4.5. B–клеточные лейкозы
| Нозология | Нормальный клеточный эквивалент | Локализация опухоли |
| Хронические лимфолейкозы | CD5+ B1–лимфоциты | Кровь |
| Острый лимфобластный лейкоз | Ранние лимфоидные клетки–предшественницы | Костный мозг и кровь |
| Пре–B–лейкоз | Пре–B–клетки | То же |
| Фолликулярная лимфома Бёркетта | Зрелые B–лимфоциты | Периферические лимфоидные органы (фолликулы) |
| Макроглобулинемия Вальденстрема | IgM–секретирующие B–лимфоциты | Периферические ткани |
| Множественная миелома (плазмоцитома) | Плазматические клетки (могут быть иммуноглобулины разных классов) | Костный мозг |
Нормальные клеточные гены, контролирующие пролиферацию, называют протоонкогенами. Онкогенами в своё время были названы открытые гены РНК–содержащих вирусов, ответственные за опухолевое перерождение инфицированных вирусом клеток.
Позже выяснили, что эти гены вирусы «списывают» с клеточных генов, которые и назвали протоонкогенами. Протоонкогеном является антиапоптозный ген bcl–2.
4.9. Конститутивные иммуноглобулины (нормальные антитела)
Если в организм не проникает ни один патоген и ни один Аг иной природы (это, конечно, возможно только в эксперименте), в крови и биологических жидкостях этого организма тем не менее будут присутствовать (и в немалом количестве) иммуноглобулины. Есть они и у естественно развивающихся организмов. Такие иммуноглобулины называют нормальными, или конститутивно синтезируемыми. Что это за иммуноглобулины? Это аутоантитела, т.е. АТ, направленные против молекул собственного организма. Они принадлежат к классам М, G и А, у взрослых большинство — к IgG. АТ полиреактивны, т.е. способны связывать множество Аг, как ауто–, так и экзогенных. В связывании Аг этими нормальными АТ, как показывают опыты по направленному мутагенезу, принимает участие область CDR3 в VH–домене тяжёлых цепей и Аг–связывающий центр кодируется зародышевыми V–генами.
Как правило, эти АТ имеют низкую аффинность, но высокую авидность к аутоантигенам. Константы диссоциации для нормальных иммуноглобулинов колеблются от 10–5 до 10–8 М. Но, например, для нормального аутоантитела к ИЛ–1a константа диссоциации составляет 5´10–11 M. Нормальные IgM–АТ обнаруживают уже в пуповинной крови новорождённых, и спектр их реактивности консервативно сохраняется во взрослой жизни. Это говорит о том, что нормальные иммуноглобулины подвержены некоему положительному отбору в онтогенезе. Выявлены следующие Аг — мишени для нормальных иммуноглобулинов: идиотопы вариабельных областей других иммуноглобулинов; каркасные и невариабельные участки молекул T–клеточных Рц для Аг; молекулы CD4, CD5 и HLA–I; Fcg–Рц; лиганды для молекул межклеточной адгезии и др.
Есть основания полагать, что нормальные АТ выполняют ряд весьма важных для здоровья организма функций:
· первая линия «обороны» против инфекций;
· удаление из организма отживших клеток и продуктов катаболизма;
· представление Аг для T–лимфоцитов (иммуноглобулины на мембране B–лимфоцитов связывают растворимые Аг в крови, лимфоцит интернализует их рецепторопосредованным эндоцитозом, и внутри клетки образуются комплексы пептидов с молекулами главного комплекса гистосовместимости, которые экспрессируются на мембране B–лимфоцита);
· поддержание гомеостаза аутоиммунной реактивности;
· противовоспалительное действие (нейтрализация суперантигенов; индукция синтеза противовоспалительных цитокинов; аттенуация комплементзависимого повреждения тканей и др.).
Например, появляются данные о том, что в случае клинической манифестации некоторых патологических аутоиммунных болезней, в патогенезе которых действуют IgG–аутоантитела, имеет место дефект регуляции этих патогенных IgG со стороны нормальных антиидиотипических IgM–АТ. Показано, что в периоды ремиссии аутоиммунных васкулитов, в патогенезе которых действуют антинейтрофильные IgG–АТ (ANCA), в крови в достаточных количествах появляются антиидиотипические по отношению к ANCA IgM–АТ. Вероятно, лечебные эффекты от введения людям с разными заболеваниями препаратов донорских иммуноглобулинов объясняются именно комплексным «противовоспалительным» действием нормальных иммуноглобулинов.
Глава 5. T–ЛИМФОЦИТЫ. ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ
5.1. Дифференцировка T–лимфоцитов
Сущность дифференцировки всякого лимфоцита, как Т, так и В, состоит в экспрессии антигенраспознающего Рц и необходимых дополнительных сервисных молекул, чтобы факт распознавания Аг имел действенные последствия, направленные на санацию организма от повреждённых тканей. Эти сервисные молекулы (мембранные, а также секретируемые цитокины) обеспечивают взаимодействие T–лимфоцитов с другими клетками организма.
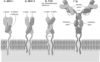
Рис. 5.1. Структура белков надсемейства иммуноглобулинов [11]. А. Молекула MHC класса I состоит из a‑цепи, внемембранная её часть связана с короткой цепью b2-микроглобулина. Б. Молекула MHC класса II состоит из двух СЕ: более длинной a‑цепи и b‑цепи. Часть каждой цепи выступает на поверхности клеточной мембраны, цепь содержит трансмембранный участок и небольшой фрагмент в цитоплазме. В. Молекула рецептора T–клеток состоит из двух цепей: a и b. Каждая цепь предcтавлена двумя внеклеточными иммуноглобулин–подобными доменами, cтабилизированными при помощи S-S cвязей, — вариабельным внеклеточным NH-концом и цитоплазматичеcким cтабильным COOH-концом. SH–группа, приcутcтвующая в цитоплазматичеcком фрагменте a‑цепи, может взаимодейcтвовать c мембранными или цитоплазматичеcкими белками. Г. Мономер молекулы IgM встраивается в плазматическую мембрану B-лимфоцитов, это рецептор Аг. Молекулярная структура рецепторов T-клеток весьма сходна со структурой молекул MHC и иммуноглобулина. Разнообразие структуры рецепторов T-лимфоцитов и иммуноглобулинов обеспечивается возможностью сайт-специфической рекомбинации множества различных генных сегментов, кодирующих отдельные фрагменты молекулы.
Антигенраспознающий Рц T–лимфоцитов обозначают TCR (T–Cell Receptor). TCR кодируется генами из надсемейства иммуноглобулинов, т.е. родственниками генов иммуноглобулинов. Молекула TCR представляет из себя по размеру и форме как бы 1/4 молекулы иммуноглобулина — аналог одного Fab–фрагмента. TCR является гетеродимером — состоит из двух равновеликих полипептидных цепей. У млекопитающих известно две разновидности пар цепей в TCR. В одной паре цепи обозначают a и b, соответствующие T–лимфоциты — Tab. Вторая пара цепей — g и d, соответствующие T–лимфоциты обозначают Tgd. Каждый индивидуальный T–лимфоцит несет какой–либо один вариант Рц — либо ab либо gd.
В отличие от иммуноглобулинов TCR исключительно трансмембранные молекулы, т.е. T–лимфоцит всегда работает собственным «клеточным телом».
Более того, в отличие от иммуноглобулинов TCRab не способен распознавать (связывать) растворимые Аг. Это относится по крайней мере к Tab и подавляющему большинству Аг, за исключением патогенных суперантигенов.
Что же тогда распознают T–лимфоциты? Природой T–лимфоциты предназначены для распознавания поверхностных структур собственных клеток организма. Если что-то на поверхности своих клеток будет «раздражать» T–лимфоцит (например, примесь вирусных пептидов), то он постарается организовать уничтожение повреждённой клетки.
T–лимфоцит распознаёт (связывается с) комплекс MHC–I или MHC–II с неким пептидом, который и есть Аг в общем понимании. Один определённый участок молекулы TCR вступает в химическую связь с молекулой MHC–I/II, второй участок того же TCR в тот же момент времени вступает в связь с пептидом–Аг. Этот феномен называют двойным распознаванием,илиMHC–рестрикцией T–лимфоцитов. Феномен был открыт Р. Цинкернагелем и П. Дохерти, в 1996 г. они получили за это открытие Нобелевскую премию. Их первая публикация в журнале «Nature» относится к 1974 г. и занимает всего половину страницы. Но это открытие многие расценивают как самое значимое в иммунологии за последние 100 лет.
Существенно, что пептид–Аг присоединяется к молекулам MHC не снаружи клетки, т.е. он не сорбируется на поверхности клетки. Пептид–Аг встраивается в комплекс с MHC при формировании конформации молекул MHC после их биосинтеза в клетке и перед их экспрессией на клеточной мембране. Молекул MHC без пептидов вообще не бывает на наружной мембране клеток, они просто не могут быть экспрессированы, не примут правильной конформации, если ещё внутри клетки не вступят в связь с пептидом определённой длины.
Таким образом, подавляющее большинство T–лимфоцитов с Рц TCRab не распознают свободных нативных Аг (ещё раз подчеркнём отличие от иммуноглобулинов, которые как раз умеют работать с нативными Аг в том виде, в каком «судьба забросила» их во внутреннюю среду организма или даже на слизистые оболочки). Другие клетки должны каким-то образом пропустить Аг через себя и выставить его на своей мембране в комплексе с MHC–I/II, чтобы T–лимфоцит «обратил на Аг своё внимание». Это и есть феномен представления Аг(или презентации Аг) T–лимфоциту. Какие клетки в организме способны быть АПК и механизмы этого процесса мы разберём в отдельном разделе, здесь лишь отметим строгую необходимость представления Аг T–лимфоциту другими клетками.
5.2. Рецептор T–лимфоцитов для антигена (TCR)
Вторичная и третичная структуры Рц TCRab и TCRgd похожи, но есть отличия: TCRgd по трёхмерной конфигурации больше похожи на иммуноглобулины. Cтроение Рц T–лимфоцитов для Аг удобнее рассмотреть на примере TCRab. T–лимфоциты с ab–Рц — это более «привычные» T–лимфоциты, которые стали известны на несколько десятков лет раньше, чем Tgd; 99% T–лимфоцитов, проходящих лимфопоэз в тимусе, — это Tab.
TCRab
TCR был открыт как особая молекула на клеточной мембране T–лимфоцитов, с которой связываются моноклональные АТ, полученные как специфично реагирующие с индивидуальным клоном T–лимфоцитов. С помощью таких АТ «увидели», что на отдельном T–лимфоците экспрессировано около 30 тыс. таких антигенспецифичных молекул. Быстро изучили молекулярную структуру Рц. Оказалось, что собственно антигенсвязывающая часть его состоит из двух равновеликих полипептидных цепей — a (мол. масса 40–60 тыс., кислый гликопротеин) и b (мол. масса 40–50 тыс., нейтральный или основный гликопротеин). Каждая из цепей имеет по два домена во внеклеточной части, гидрофобную положительно заряженную (за счёт остатков лизина и аргинина) трансмембранную часть и короткий (из 5–12 остатков АК) цитоплазматический участок. Между собой цепи соединены одной дисульфидной связью выше мембраны. Каждый из внеклеточных доменов имеет по одному сайту (месту связывания) гликозилирования и соответственно гликозилирован.
Наружные внеклеточные домены обеих цепей имеют вариабельный АК–состав от Рц к Рц (от лимфоцита к лимфоциту), гомологичные V–области молекул иммуноглобулинов и названные одноимённо — V–областью TCR. Проксимальные домены обеих цепей гомологичны константным областям молекул иммуноглобулинов и также названы одноимённо — C–областью TCR. Соответственно обозначены и гены, кодирующие эти области полипептидов: Va и Сa, Vb и Сb.
Дата добавления: 2016-07-18; просмотров: 2572;
Поиск по сайту
Узнать еще
- a-спираль b-складчатая структура
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
