СТРУКТУРА И ФУНКЦИИ ИММУННОЙ СИСТЕМЫ 11 глава
Именно V–области a– и b–цепей вступают в связь с комплексом MHC–I/II — пептид. Место контакта с пептидом в a–цепи — участок CDR3, причём с центральной частью пептида вступает в связь участок контакта Va–Ja.
Короткий цитоплазматический участок как у a-, так и b–цепи физически недостаточен для обеспечения проведения сигнала внутрь клетки. Для проведения сигнала с TCR служат входящие в рецепторный комплекс ещё 6 инвариантных (т.е. одинаковых у всех T–лимфоцитов) полипептидных цепей: g , d, две e и две z. Комплекс a + b + 2e, как выяснили, и составляет мембранную структуру CD3, идентифицированную раньше по реактивности с моноклональными АТ, связывающими все без исключения T–лимфоциты. Цепи g, d и e имеют внеклеточный, трансмембранный (отрицательно заряженный за счёт остатков аспарагина и поэтому электростатически связанный с трансмембранными участками a- и b–цепей) и цитоплазматический участки. В случае генетических дефектов в комплексе CD3 a– и b–цепи на мембране T–лимфоцита не появляются. Это свидетельствует о том, что полипептиды g, d и e необходимы ещё и для формирования правильной конформации a– и b–цепей и экспрессии их на мембране.
Полипептиды комплекса CD3 — g, d и e высокогомологичны друг другу. Их гены локализованы рядом, все в хромосоме 11 у человека, и, очевидно, произошли путём дупликации от одного предкового гена. Две z–цепи соединены между собой дисульфидной связью, в мембране встроены между трансмембранными участками a- и b–цепей и бoльшая часть полипептидной z–цепи уходит в цитоплазму клетки. Там эта цепь и «задействуется» в реакциях проведения сигнала при связывании Аг V–областями a и b на внутриклеточные структуры и процессы.
Во всех цитоплазматических участках полипептидных цепей g, d, e и z содержатся последовательности АК — ITAM, которыми эти полипептиды взаимодействуют с протеинтирозинкиназами цитозоля (активация этих ферментов и составляет начало биохимических реакций по проведению сигнала).
Схематически структура TCR, состоящего из 8 или 10 (по одной или две пары a + b на проводящий комплект 2e + d + g+ 2z) полипептидных цепей, представлена на рис. 5.1.
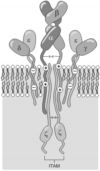
Рис. 5.2. Строение рецептора T–лимфоцитов для антигена (TCRab) (схема). Антигенсвязывающая область Рц формируется a- и b–цепями; цепи g, d, e (вместе их называют комплексом CD3) необходимы для экспрессии a- и b–цепей, их стабилизации и, вероятно, проведения сигнала внутрь клетки; z–цепь, самая «внутриклеточная», обеспечивает проведение сигнала внутрь клетки.
Рц для Аг присоединяет свои лиганды ионными, водородными, ван–дер–ваальсовыми и гидрофобными связями. Рц достаточно «жив» и подвижен. При связывании с Аг его конформация существенно изменяется. Как показывают исследования с применением самых современных методов с использованием трансгенных по TCR мышей (у которых все T–лимфоциты имеют единственный вариант TCR) и коллекции синтетических пептидов, а также контролируемых комплексов пептидов с растворимыми молекулами MHC, один TCR потенциально способен связывать достаточно много разных Аг (если под Аг понимать 9–членные пептиды): по минимальным оценкам 103, по максимальным — 107, по наиболее точным оценкам порядка 105. Причём один TCR связывает не только родственные по структуре Аг (их понимают как перекрестно реагирующие), но и Аг, не имеющие гомологии в структуре.
5.3. Гены a- и b–цепей рецепторов T–лимфоцитов для антигена
Гены a- и b–цепей (а также g и d–цепей) устроены гомологично генам иммуноглобулинов и претерпевают соматическую рекомбинацию (перестройку ДНК) в процессе дифференцировки T–лимфоцитов. Это и обеспечивает генерацию разнообразия антигенсвязывающих свойств TCR, оцениваемого так же, как и для иммуноглобулинов: теоретически порядка 1016–1018 вариантов, реально порядка 109 (соответственно числу лимфоцитов в организме).
Гены a–цепи имеют 70–80 V–сегментов и 61 J–сегмент, комбинации которых по одному из каждой области обеспечивают дополнительное разнообразие антигенсвязывающих свойств TCR. Гены b–цепи имеют 52 V–сегмента, 2 D–сегмента и 13 J–сегментов, а также 2 C–сегмента, расположенных, как показано на рис. 5.2. Между V– и J–сегментами a–цепи находится локус генов d–цепи второго варианта TCR — gd. Локус d состоит из трёх D–сегментов, трёх J–сегментов, одного C–сегмента. V–сегменты d–цепи (ориентировочно их всего 4) «вкраплены» среди V–сегментов a–цепи.

Рис. 5.3. Структура генов a- и b–цепей рецептора T–лимфоцитов для антигена.
Рекомбинация ДНК происходит только при объединении V–, D– и J–сегментов и катализируется тем же комплексом рекомбиназ, инициируемых продуктами генов RAG1 и RAG2, которые действуют при дифференцировке B–лимфоцитов. Только субстрат для рекомбиназ — гены TCR, а не гены иммуноглобулинов в T–лимфоцитах — определяют тканеспецифичные промоторы. Рекомбиназы «узнают» определённые гептамерные или нонамерные последовательности, фланкирующие каждый из сегментов V, J, D и называемые rss (recombination signal sequences) — последовательностями, сигнальными для рекомбинации. Перед каждым V–экзоном имеется L–экзон (от leader sequence — лидерная последовательность), который обязательно транслируется и обеспечивает правильное расположение полипептидной цепи в эндоплазматическом ретикулуме и правильную экспрессию на клеточной мембране (рис. 5.3).
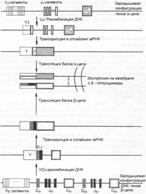
Рис. 5.4. Перестройка и экспрессия генов рецептора T–лимфоцитов для антигена (TCR).
После перестройки VJ в генах a–цепи и VDJ в генах b–цепи, а также присоединения некодируемых N– и P–нуклеотидов с ДНК транслируется РНК. Объединение с C–сегментом и удаление лишних неиспользуемых J–сегментов осуществляются при сплайсинге первичного транскрипта.
Гены a–цепи могут перестраиваться неоднократно при уже правильно перестроенных и экспрессированных генах b–цепи. Параметры разнообразия антигенсвязывающих участков T–клеточных Рц приведены в табл. 5.1.
Таблица 5.1. Параметры разнообразия антигенсвязывающих областей TCR (Рц T–лимфоцитов для антигена)
| Генетический элемент | Число вариантов | |
| | | |
| Зародышевое число V–сегментов | | |
| Зародышевое число D–сегментов | ||
| Транскрипция D–сегментов со смещением рамки считывания | — | Часто |
| Зародышевое число J–сегментов | | |
| Присоединение N– и P–нуклеотидов |
Программа для соматического гипермутагенеза в генах TCR отсутствует (в отличие от генов иммуноглобулинов).
Столь большое число — от 1018 до 109 — потенциально возможных вариантов антигенсвязывающих Рц на лимфоцитах является попыткой медленно размножающихся и медленно эволюционирующих видов многоклеточных организмов «угнаться» за изменчивостью сверхбыстро размножающихся и эволюционирующих инфекционных микроорганизмов. Природа сделала такой шаг — наделила суперизменчивостью в процессе онтогенеза в рамках одного тела единственный гистотип клеток (только лимфоциты) и единственную категорию молекул в этих клетках (антигенраспознающие Рц — TCR, иммуноглобулины). Лимфоцитарный иммунитет есть у очень небольшого числа видов земных многоклеточных животных, напомним — всего у 1,5%. Следовательно, 98,5% видов многоклеточных не имеют лимфоцитарной иммунной системы. Лимфоцитарный иммунитет понадобился для спасения от инфекций особей таких видов животных, которые оставляют единичное потомство — рожают редко и мало детенышей, значит, если не каждому, то большинству надо выжить, чтобы сохранился вид. Лимфоциты со своими Рц появились, возможно, 300 млн лет назад (вероятно, у челюстных рыб). При изучении структуруы генов RAG1 и RAG2, продукты которых начинают перестройку ДНК в генах иммуноглобулинов и TCR, было установлено, что эти уникальные гены RAG похожи на транспозоны. Возникла гипотеза, что гены RAG — ретровирусы по происхождению. Таким образом, возможно именно ретровирусная инфекция древнейших челюстных рыб привнесла новое свойство — соматическую рекомбинацию ДНК. Это свойство оказалось полезным для выживания и закрепилось отбором в лимфоцитах всех вышестоящих видов животных. Так, возможно, и возник лимфоцитарный иммунитет.
5.4. Корецепторные молекулы T–лимфоцитов — CD4 и CD8
TCRab распознаёт Аг–пептид только в связи с молекулами MHC–I или II. Каждый конкретный T–лимфоцит предназначен для работы с молекулами либо класса I, либо класса II MHC. Собственно в связь с MHC (ионными, ван–дер–ваальсовыми, водородными и гидрофобными связями) вступают вариабельные участки полипептидных цепей a и b. Но со стороны T–лимфоцита есть ещё одна из двух (у индивидуального лимфоцита какая-то одна) мембранных молекул, вступающих в связь с молекулами MHC на АПК или клетках–мишенях для T–лимфоцитов — это молекулы CD4 и CD8. Молекула CD4 имеет химическое сродство и вступает в связь с инвариантной частью молекулы MHC–II (b2–доменом), молекула CD8 — с инвариантной частью молекулы MHC–I (a3–доменом). Поэтому молекулы CD4 и CD8 называют корецепторными молекулами T–лимфоцитов. В экспериментах, моделирующих отсутствие на клетке этих молекул, требуются минимум в 100 раз большие дозы Аг, чтобы активировать T–лимфоцит.
Молекула CD4 — полипептидная цепь с молекулярной массой 55 тыс. Она имеет 4 домена во внеклеточной части и по первичной структуре принадлежит к надсемейству иммуноглобулинов. Цитоплазматический участок молекулы CD4 связан с тирозинкиназой Lck и, вероятно, вносит свой вклад в проведение сигнала внутрь клетки при состоявшемся взаимодействии Рц с лигандом. В процессе активации T–лимфоцита одну молекулу TCR «обслуживают» две молекулы CD4 (происходит димеризация молекул CD4) (рис. 5.4).
Молекула CD8 — гетеродимер из двух цепей, традиционно обозначаемых a и b. В некоторых случаях на клетке обнаруживают гомодимер из двух a–цепей. Такой гомодимер также имеет свойство взаимодействовать с молекулами MHC–I. Во внеклеточной части каждая из a- и b–цепей имеют по одному иммуноглобулинподобному домену. Между собой цепи соединены дисульфидной связью. Цитоплазматические участки цепей CD8 также связаны с тирозинкиназой Lck (см. рис. 5.4).
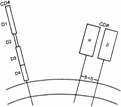
Рис. 5.5. Структура молекул корецепторов T–лимфоцитов — CD4 и CD8 (схема). Молекула CD4 — полипептидная цепь, имеюшая 4 внеклеточных домена. При вовлечении в функциональный процесс молекула CD4, вероятно, димеризуется. Молекула CD8 — гетеродимер из a– и b–цепей.
Почти все лимфоциты (99%), дифференцирующиеся на территории тимуса в постнатальной жизни, — это T–лимфоциты с TCRab; менее 1% — T–лимфоциты с TCRgd. В периферических тканях содержание Tgd больше 1% и составляет, возможно, 10–50% всех T–лимфоцитов организма. Но дифференцируется большинство Tgd не в тимусе, а в слизистых оболочках, в первую очередь ЖКТ.
5.5. Дифференцировка T–лимфоцитов в тимусе. Позитивная и негативная селекция тимоцитов
Процессы дифференцировки, происходящие в тимусе, изучены достаточно подробно и представляют следующую последовательность событий.
· Тимоциты дифференцируются из общей стволовой кроветворной клетки. На клетках–предшественницах T–лимфоцитов ещё вне тимуса у человека обнаружены такие мембранные маркёры, как CD7, CD2, CD34 и цитоплазматическая форма CD3.
· Коммитированная к дифференцировке в T–лимфоцит клетка–предшественница мигрирует из костного мозга через стенку венул в кортико-медуллярной области и оттуда перемещается в субкапсулярную зону коры тимуса. Здесь происходит медленная пролиферация клеток — примерно в течение 1 нед. Характерной особенностью лимфоидных клеток–предшественниц является сохранение в них экспрессии уникального фермента стволовой кроветворной клетки — теломеразы, репарирующей хромосомы. Это указывает на то, что лимфоциту на его жизненном пути предстоит ещё не раз пройти циклы пролиферации. Клетки–предшественницы, коммитированные к дифференцировке в другие ростки кроветворения, утрачивают экспрессию теломеразы необратимо. Из предшественников лимфоидных клеток будут развиваться Tab (большинство), Tgd и дендритные клетки тимуса. На тимоцитах, самыми ранними известными маркёрами которых являются молекулы CD2 и CD7, в этот период экспрессированы ещё такие мембранные молекулы, как CD44 и CD25.
· Затем на клетках исчезают молекулы CD44 и CD25, клетки физически перемещаются несколько в глубь коры тимуса. В них начинаются перестройка генов b–цепи TCR и экспрессия полипептидного комплекса CD3. Экспрессируется инвариантная суррогатная полипептидная цепь, называемая pTa (р — от partner — партнёр, мол. масса 33 кД), которая заменяет собой настоящую a–цепь и позволяет перестроенной b–цепи в комплексе с CD3 оказаться экспрессированной на наружной клеточной мембране. На данной стадии деградирует белок RAG2 и ингибируется транскрипция с генов RAG1 и RAG2. Клетки активно пролиферируют и микроскопически выглядят как бласты. При этом накапливается масса клеток с уже готовой b–цепью, но с ещё неперестроенными генами a–цепи, что внесет свой вклад в будущее разнообразие a–b–гетеродимеров.
В это же время экспрессируются совместно молекулы CD4 и CD8 (по этой причине тимоциты на этой стадии развития называютдважды позитивными— CD4+/ CD8+).
· На следующем этапе клетки перестают делиться, синтезируют RAG1 и RAG2 и перестраивают Va–гены, причём несколько раз в течение 3–4 сут. Каждый вариант TCR с разной a–цепью клетка экспрессирует и «пробуется» на связывание с комплексами «пептид–MHC», экспрессированными на мембранах эпителиальных клеток тимуса. Обратим внимание на то, что пептиды в данном случае свои, т.е. являются продуктами катаболизма собственных белков АПК. Тимоциты, которые свяжут какой–либо из комплексов «пептид–MHC» с правильной (т.е. средней по силе) аффинностью, получают сигнал на выживание и продолжат дифференцировку. Тимоциты, которые не свяжут никакой из доступных комплексов «пептид–MHC», не получат сигнала на выживание, и в них будет инициирована программа на апоптоз, они погибнут. Это и называют позитивной селекцией тимоцитов. В результате позитивной селекции в тимусе погибает около 90% тимоцитов.
Если какой-то TCR связывает какой-то комплекс «пептид–MHC» со слишком высокой аффинностью, то тимоцит также может погибнуть. Это называют негативной селекцией тимоцитов. В количественном отношении негативная селекция элиминирует существенно меньше тимоцитов, чем позитивная — от 10 до 70% клеток, которые прошли позитивную селекцию.
Неоднократная перестройка и экспрессия генов a–цепи при одной и той же b–цепи в одном тимоците оставляет шанс на то, что одна клетка может нести более одного варианта TCR. По крайней мере на моделях мышей, трансгенных по генам b и a–цепей, закономерно обнаруживают второй вариант Рц на T–лимфоцитах с эндогенной a–цепью.
На короткое время с мембраны тимоцитов исчезают обе молекулы корецепторов CD4 и CD8, затем экспрессируется одна из этих молекул: на тимоцитах, TCR которых имеет сродство к молекулам MHC–I, экспрессируется корецептор CD8, и на периферии эти T–лимфоциты на всю свою жизнь будут иметь «паспортную» характеристику CD8+, или Т8. На тимоцитах, TCR которых имеет сродство к MHC–II, экспрессируется корецептор CD4, и на периферии такие T–лимфоциты будут CD4+, или Т4. Есть данные, полученные на экспериментальных моделях на мышах, показывающие, что один и тот же TCR (трансгенный), экспрессированный в паре с CD4, связывает пептид–Аг в комплексе с MHC–II, а экспрессированный в паре с CD8 распознаёт другой пептид–Аг в комплексе с MHC–I.
Анатомически процессы селекции проходят ближе к границе коркового и мозгового слоев тимуса при взаимодействии развивающихся тимоцитов с эпителиальными клетками стромы тимуса и с дендритными клетками костномозгового происхождения. На клетках эпителия тимуса экспрессированы молекулы адгезии LFA–3 и ICAM–1, а на тимоцитах — комплементарные им молекулы адгезии соответственно CD2 и LFA–1. Эти адгезивные взаимодействия удерживают тимоциты на территории тимуса на необходимый период времени. На эпителиальных и дендритных клетках мозговой зоны и на внешних структурах телец Гассаля (телец тимуса) экспрессирована в большом количестве мембранная молекула CD30L (CD30–лиганд). На тимоцитах же, активированных аутоантигенами, экспрессируется молекула CD30. Полагают, что именно взаимодействие CD30L с CD30 между стромой и тимоцитами, имеющими на мембране набор молекул CD4+CD8+CD45R0+CD30+ИЛ–4R+, обеспечивает сигнал на апоптоз для тимоцитов и это и есть негативная селекция аутореактивных клонов тимоцитов.
В кратком виде этапы дифференцировки T–лимфоцитов в тимусе приведены в табл. 5.2. Дифференцировка T–лимфоцитов в тимусе продолжается около 3 нед.
Таблица 5.2. Этапы дифференцировки T–лимфоцитов в тимусе
| Анатомическая зона в тимусе | Мембранный фенотип тимоцитов и их клеточное окружение | Основные процессы, происходящие с тимоцитами |
| Кора периферической зоны | CD3–/CD4–/CD8– Эпителиальные клетки коры | Интенсивная пролиферация (митозы) дважды негативных клеток–предшественниц |
| TCR– | Экспрессия | |
| Глубокие слои коры | TCR– | Дважды позитивные (CD4+ и CD8+) тимоциты с экспрессированной |
| Граница коркового и мозгового вещества | TCR– | Дважды позитивные тимоциты с полноценным |
| Мозговое вещество | TCR– | Однопозитивные (либо CD4+, либо CD8+) зрелые неиммунные T–лимфоциты |
Рассмотрим некоторые подробности дифференцировки T–лимфоцитов в тимусе.
Tab и Tgd происходят из общей клетки–предшественницы. В развивающемся тимоците практически одновременно начинается перестройка генов b, g и d–цепей. Если обе g и d–цепи успеют перестроиться «in–frame» (т.е. получится такая ДНК, с которой возможна транскрипция РНК), то лимфоцит будет T gd. Но если первой успеет перестроиться b–цепь и экспрессироваться на мембране в паре с pTa, то это становится сигналом к запрету на продолжение перестройки генов g и d, а также сигналом к пролиферации клетки, экспрессии корецепторных молекул CD4 и CD8 и чуть позже к перестройке генов и экспрессии a–цепи. При перестройке генов a–цепи происходит необратимая делеция d–локуса, который расположен внутри генов a–цепи. Непродуктивная с первой попытки перестройка генов b–цепи не означает, что тимоцит обречен на гибель. Структура генов b–цепи (см. рис. 5.2) позволяет предпринять ещё одну попытку, и продуктивность перестройки генов b–цепи превышает 80%.
Вероятный лиганд для комплекса pTa–b на мембране тимоцитов — молекула CD81, экспрессированная на эпителиальных клетках стромы тимуса.
Перестройка генов a–цепи происходит неоднократно, пока не получится продуктивный вариант, причём без аллельного исключения (т.е. параллельно на обеих гомологичных хромосомах). До этапа позитивной селекции на тимоцитах могут быть экспрессированы два варианта TCR с одинаковыми b, но разными a–цепями. Позитивная селекция оставит единственный вариант Рц, но иногда может быть и не единственный.
Позитивная селекцияна удовлетворительное связывание с комплексами своих пептидов со своими молекулами MHC происходит на стадии развития тимоцитов с низкой плотностью экспрессии TCR и совместной экспрессией корецепторов CD4 и CD8. Клетки–партнёры тимоцитов в данном процессе, клетки–носители молекул MHC — это эпителиальные клетки стромы коры тимуса. Эпителиальные клетки тимуса продуцируют такие цитокины, как интерлейкины 1, 3, 6 и 7, LIF, GM–CSF. Они же, эпителиальные клетки тимуса, являются продуцентами внутритимического стероидного гормона, а именно прегненолона, участие которого необходимо в регуляции экспрессии генов в дифференцирующихся тимоцитах, в том числе в индукции апоптоза тимоцитов при позитивной и негативной селекции. (В эксперименте, если мыши ввести фармакологические дозы глюкокортикоидных гормонов, то за несколько часов произойдет «глюкокортикоидная тимэктомия» — более 99% тимоцитов погибнет апоптозом, от органа останется небольшая «плёночка», содержащая немного стероидрезистентных T–лимфоцитов).
В качестве примера экспериментов, ставших источником этих знаний, приведём опыты с мышами — радиационными костномозговыми химерами. Если летально облученным реципиентам одной из родительских линий, например гаплотипа MHCA, трансплантировать клетки костного мозга интактных доноров-гибридов MHCА´B, то дифференцированные в тимусе реципиента (MHCA) T–лимфоциты будут способны распознавать Аг только в комплексе с молекулами MHCA, но не MHCB.
«Профессиональные» АПК периферии — это клетки костномозгового происхождения, а именно дендритные клетки, B–лимфоциты и моноциты/макрофаги. Это поможет понять результаты следующего опыта. Если летально облученным мышам–реципиентам гаплотипа MHCВ трансплантировать костный мозг от доноров гаплотипа MHCA, то дифференцирующиеся в организме (в тимусе) реципиента T–лимфоциты оказываются неспособными развивать иммунный ответ в периферической лимфоидной ткани. В тимусе их «отобрали» на распознавание MHCВ, а представляют Аг им в организме АПК генотипа MHCA, т.е. «своего родного» для T–лимфоцитов, но они его «не приучены» распознавать: иммунорецепторы вообще распознают не «своё» и «чужое». Что будут, а что не будут распознавать антигенраспознающие Рц лимфоцитов, обусловлено не столько генетической программой, сколько конкретными ситуациями, возникающими в процессе онтогенеза.
Описанный выше эксперимент показывает, почему при трансплантации костного мозга у людей с лечебной целью строго необходимо совпадение по MHC: не столько для избежания отторжения трансплантата реципиентом, сколько для обеспечения возможности дифференцировки иммунокомпетентных T–лимфоцитов из стволовых клеток костного мозга донора в тимусе реципиента.
У мышей, трансгенных по перестроенному гену TCR, распознающему пептид в комплексе с MHC–I, все T–лимфоциты будут нести только этот трансгенный Рц и все T–лимфоциты в организме будут только CD8+. Если трансгенный TCR комплементарен Аг в комплексе с MHC–II, то все T–лимфоциты в организме такой мыши будут только CD4+.
Та же закономерная зависимость распознавательной ориентировки TCR либо на MHC–I, либо на MHC–II и экспрессии на зрелом лимфоците соответственно или CD8, или CD4 проявляется у людей с наследственным иммунодефицитным синдромом «голых лимфоцитов». Есть варианты этого синдрома, когда генетический дефект проявляется в отсутствие экспрессии на эпителиальных клетках тимуса и на лейкоцитах молекул MHC–I. У таких пациентов не обнаруживают CD8+ T–лимфоцитов. При вариантах синдрома с отсутствием экспрессии на клетках MHC–II у больных в периферических лимфоидных органах нет T–лимфоцитов CD4+.
Корреляция экспрессии TCR с молекулами CD8 и CD4 в описанном варианте сильная, но не абсолютная. В организме есть и иные субпопуляции T–лимфоцитов, например TCRab/CD4–/CD8–, TCRaa/CD8aa (молекула CD8 состоит из двух одинаковых a–цепей, т.е. гомодимер, а не гетеродимер из одной a– и одной b–цепи, как в «классическом» варианте молекулы CD8). Кстати, T–лимфоциты именно с таким фенотипом мембраны являются преобладающей субпопуляцией среди внутриэпителиальных лимфоцитов слизистой оболочки ЖКТ. Не зависит от CD4 и CD8 и экспрессия TCRgd.
T–лимфоциты, экспрессирующие CD4, и T–лимфоциты, экспрессирующие CD8, предназначены для выполнения разных функций в предстоящих иммунных ответах. Но во всех случаях независимости TCR от CD4 или CD8 лимфоциты с такими TCR распознают Аг независимо от MHC–I/II. Правда, это не означает (по крайней мере не всегда очевидно) независимость распознавания Аг от других молекул, способных представлять Аг, но не являющихся ни MHC–I, ни MHC–II. Примером таких антигенпредставляющих молекул является CD1. Если молекулы MHC–I и II «работают» (т.е. формируют комплексы) исключительно с пептидными Аг, то «неклассические молекулы MHC» специализируются на комплек-сообразовании с Аг другой химической природы — небелковой (углеводными, липидными, фосфатсодержащими). С молекулами кластера CD1 взаимодействуют особые субпопуляции зрелых T–лимфоцитов, а именно «дважды негативные» TCRab+/CD3+/CD4–/CD8– и особая субпопуляция TCRgd+/CD3+/CD8+.
Процессы негативной селекции тимоцитов идут при взаимодействии их с профессиональными АПК — дендритными и макрофагами, что иллюстрируют эксперименты с костномозговыми радиационными химерами мышей. Если реципиентов любой из родительских линий (MHCA или MHCB) летально облучить и трансплантировать им костный мозг от гибридов MHCА´B, то в тимусе таких мышей будут развиваться T–лимфоциты, не отторгающие кожный лоскут ни от донора MHCA, ни от донора MHCB, т.е. развивается толерантность к Аг как реципиента, так и донора, которые в тимусе могут быть представлены только на тимоцитах, дендритных клетках и макрофагах. При некоторых комбинациях линий мышей в таких экспериментах кожный трансплантат, однако, отторгается. Это объясняют тем, что для отторжения ткани существенны и неглавные Аг гистосовместимости(англ. minor histocompatibility antigens), которые в данном случае присутствуют в ткани донорской кожи, но не присутствуют на донорских АПК в тимусе и на уровне дифференцировки лимфоцитов в тимусе толерантность к ним не возникает. Неглавные Аг гистосовместимости при отторжении трансплантатов у людей иногда имеют большее значение, чем Аг главного комплекса гистосовместимости, что будет проиллюстрировано в соответствующем разделе.
Созревшие в тимусе T–лимфоциты мигрируют в T–зоны периферических лимфоидных органов. Путей выхода тимоцитов из тимуса на основании гистологических наблюдений можно предполагать три: а) свободный — через эфферентный лимфатический сосуд в кортико-медуллярной зоне; б) путём экстравазации через стенку посткапиллярных венул с высоким эндотелием там же в кортико-медуллярной зоне; в) путём экстравазации через стенку капилляров в русле транскапсулярных артерий.
Базовым свойством лимфоцитов является самораспознавание в смысле распознавания клеток своего организма. Теперь это можно понять несколько лучше: дифференцировка и отбор антигенраспознающих Рц T–лимфоцитов в тимусе идут при взаимодействии (комплементарном связывании) исключительно с эндогенными, т.е. со своими молекулами MHC в комплексе со своими, но разнообразными пептидами. Приведём совпадающее с нашим мнение авторитетного экспериментатора и теоретика Ч. Джейнвэя и его коллег. Опыты на мышах, трансгенных по гену b–цепи TCR, а также с нокаутом по определённому гену MHC (Н–2Ма) в разных аранжировках привели этих авторов к заключению (гипотезе, но обоснованной), что не только в тимусе в процессе лимфопоэза идёт отбор T–лимфоцитов на способность правильно связывать своим TCR комплексы «свои пептиды — MHC», но и на периферии жизнеспособность T–лимфоцитов в течение длительного времени поддерживается сигналами на выживание от тех же комплексов «свои пептиды — MHC», экспрессированных на дендритных клетках в T–зависимых зонах периферических лимфоидных органов. В отсутствие такого сигнала T–лимфоцит быстро погибает. Более того, такой свой сигнал поддерживает некий «базальный» уровень активации и, возможно, пролиферации жизнеспособных T–лимфоцитов на периферии в течение всей жизни организма. Но эта пролиферация отличается от экспансии клона лимфоцитов в процессе развития активного продуктивного иммунного ответа на Аг, подлежащий элиминации из организма. Пролиферация по сигналу от своего в норме не сопряжена с синтезами и секрецией активных цитокинов T–лимфоцитов и индукцией иммунного воспаления. Таким образом можно объяснить и понять то реально существующее явление, что после 15-летнего возраста, когда T–лимфопоэз в тимусе существенно угасает, тем не менее в течение всей взрослой жизни в организме есть, причём достаточно постоянное (не снижающееся прогредиентно с возрастом) количество периферических Tab–лимфоцитов.
Дата добавления: 2016-07-18; просмотров: 2431;
Поиск по сайту
Узнать еще
- a-спираль b-складчатая структура
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
