Ритмы роста растений
Многие процессы у растений (рост, развитие, обмен веществ и т. д.) имеют ритмический характер. О многих явлениях, которые происходят с правильной периодичностью во времени и пространстве, говорят, что им присущ ритм. Когда периодичность сохраняется и в отсутствии внешних воздействий, ритмы называют эндогенными. Примером могут служить «суточные» движения, а именно, ночное опускание листьев фасоли, приподнятых в дневное время, раскрывание и закрывание цветков каланхоэ, ритмические суточные изменения скорости роста колеоптилей овса.
Вот такая эндогенно-направленная суточная ритмика называется циркадной (латинского цирка – около, диес – день). Клеточный механизм, генерирующий ритм, обычно называют биологическими часами; положение органа (например, листа) в любой момент цикла показывает «циркадное время» (время контролируемое внутренним осциллятором). Циркадные колебания такого типа встречаются во всех эукариотичных организмов, включая микроорганизмы, растения и животных.
Чтобы выявить природу осциллятора, были сделаны попытки установить, какие условия могут изменить период (время) или фазу ритма. Эти опыты (механические воздействия, изменение освещения, температуры, ионы Li+, тяжелая вода, этанол) дали возможность сделать вывод, что периодические изменения свойств мембраны являются составной частью «часового» механизма.
Очень интересным фактом является независимость продолжительности периода в широких интервалах температуры. С функциональной точки зрения это неудивительно, так как любые часы были бы плохими показателями времени, если бы скорость их хода зависела от температуры.
Сегодня предлагают несколько механизмов биологической компенсации температуры. Например, считают, что происходят два противоположных процесса: один из которых при повышении температуры ускоряется, а другой – замедляется. Другая гипотеза связана с изменением длины цепи и степени насыщаемости мембранных липидов.
Кроме суточной периодичности рост растений имеет и сезонную периодичность. Она может быть продемонстрирована на примере образования годичных колец в древесине растений умеренного пояса, у которых рост стволов в толщину достигает максимума в летний период и прекращается осенью.
С другой стороны, внешние воздействия могут вызвать движение органов, т. е. растением свойственна раздражимость (возбудимость). Восприятие возбуждения – сложный процесс, включающий физический этап (например, поглощение света пигментом) и возникновение электрической активности, например в виде потенциала действия (ПД).
Движение – одно из наиболее заметных проявлений жизни даже в том случае, если оно совершается медленно, как это происходит у растений. У растений наблюдаются главным образом движения органов путем изгиба, скручивания и т. д.
В процессе роста и развития растение изменяет свое положение в пространстве. Развертывание цветочной почки при распускании цветка мы рассматриваем как ростовой процесс, но если тот же цветок закрывается вечером и вновь раскрывается утром, то мы говорим уже о движении, несмотря на общность механизмов, лежащих в основе обоих явлений.
Движение растений – это положения органов растений в пространстве, обусловленное разными факторами среды: светом, температурой, силой тяжести, химическими агентами и др.
Обычно выделяют четыре типа раздражителей: механические, физические, химические и повреждения.
Механические раздражения подразделяют на сотрясение, повторяющуюся деформацию растения или его части, и прикосновение, т. е. местное давление.
Физическое раздражение. Целесообразно выделить электростимуляцию, температуру, фотораздражение. Сюда можно отнести и ионизирующие излучения, т. е. воздействие любых электромагнитных излучений, условно исключая при этом воздействие видимым светом, как наиболее адекватный случай действия излучения вообще.
Электростимуляция хотя и не является естественной, но она адекватна природе биоэлектрического ответа, т. к. непосредственно связана с переносом заряда, обеспечивающего снижение Ψм на пороговую величину.
Температурный фактор может быть весьма эффективным раздражителем растений, удобным для применения в электрофизиологическом эксперименте. Этот фактор широко использовал Дж. Бос, считая его «идеальным способом раздражения», т. к. его можно применять в заданных количествах и без какого-либо повреждения много раз подряд.
Изменение интенсивности видимого света (фотораздражение) адекватный раздражитель для фотосинтезирующих зеленых растений.
Действие ионизирующих излучений исследованы очень слабо, хотя и отмечены потенциалы действия (ПД) при их действии. Неясно является ли пороговая деполяризация в этих случаях прямым следствием действия излучения или она передается, как и при фотостимуляции, от внутриклеточных пигментных систем. Последнее подтверждается авторами, показавшими, что изменение электрического потенциала клетки нителла при рентгеновском облучении зависит от того, были ли клетки в темноте или адаптировались к освещению.
Химическое раздражение. Можно выделить несколько групп химических раздражителей, вызывающих ПД у растений: ионную стимуляцию, гидростимуляцию и раздражение физиологически активными веществами.
Ионная стимуляция. Наиболее распространенным раздражителем, вызывающим ПД у высших растений является раствор KCl. Чаще всего используется 1М раствор KCl, но ПД, возникают и при действии 0,1 М и даже 0,01 М. Это свидетельствует о том, что пороговая деполяризация, вызывающая ПД, может осуществляться при физиологически умеренных изменениях концентрации K+ вне клеток. Используются в качестве раздражителей и другие соли (0,5–3 М NaCl).
Гидростимуляция: увлажнение слегка подсохших корней является эффективным для вызова ПД в стебле.
Физиологически активные вещества. Сюда относятся очень разнообразная группа раздражителей естественного и искусственного происхождения, способных приводить к генерации ПД. Например, 2,4-Д (дихлорфеноксиуксусная кислота) в концентрации 10–1–10–2 М, ауксины и др. Однако, химические раздражители, как правило, приводят к нарушению процессов возбуждения, поэтому в последнее время они не примеряются.
Повреждениявызывают несколько иную генерацию электрической активности, а именно возникновение вариабильного потенциала (ВП).
Терминология, которой пользуются в физиологии раздражения (такие понятия, как раздражение, возбуждение, реакция) заимствована из нейрофизиологии. В настоящее время восприятие возбуждения принято обозначать термином перцепция (рецепция).
Во многих случаях, как уже отмечено, восприятие раздражителя включает возбуждение. Возбуждение представляет собой изменение состояния клетки; оно начинается с изменения электрического потенциала и последующего возникновения ПД, что приводит к временному отсутствию возбудимости (восприятие раздражения). В этом случае говорят о рефрактерном периоде: абсолютном и относительном.
Механизмы возникновения и распространения ПД особенно хорошо изучен на гигантских клетках междоузлий харовых водорослей (рис. 6.19).
|
 |
Он возникает под действием различных раздражителей вызывающих вход ионов свободного Са2+ в цитоплазму из внешней среды или из внутриклеточных компартментов. Повышение концентрации свободного Са2+ в цитоплазму приводит к активации хлорных каналов на плазмалемме и тонопласте, т. е. к выходу из цитоплазмы ионов Cl-. Реполяризация осуществляется за счет входа в цитоплазму ионов K+.
Аналогичен механизм возникновения ПД у высших растений. Особенностью является участие Н+-АТФазной помпы в восстановлении ионного внутриклеточного гомеостаза.
Движение отдельных органов растений при действии различных раздражителей разделяют на два больших класса: тропизмы и настии.
Тропизмы(от греческого тропос – поворот) – процессы изгибания растущих частей прикрепленных растений, вызываемые односторонне действующими раздражителями (свет, сила тяжести и др.). Тропизмы являются результатом более быстрого роста клеток на одной стороне побега, корня или листа. При характеристике тропизмов сначала называют вызывающий их фактор. Так фототропизм – ориентировка осевых органов растений (стебель, корень, лист) к одностороннему освещению, которая выражается в направленном росте или изгибе к свету (положительный тропизм), или от света (отрицательный тропизм).
Родоначальником учения о фототропизме считают Ч. Дарвина. Он выяснил, что световое раздражение воспринимается верхушкой, ответная реакция (изгиб) возникает в зоне растяжения, лежащей ниже. Таким образом, клетки, воспринимающие световой сигнал (рецепторы), и клетки, отвечающие на раздражение, пространственно разобщены. Это позволило предположить существование веществ, которые синтезируются в одном месте, а действуют в другом (гормоны).
В 1928 г. физиологи Н. Г. Холодный и Ф. В. Вент независимо друг от друга сформулировали гормональную теорию фототропизма. В основе этой теории лежит представление о перераспределении гормонов в верхушке побега. Одностороннее освещение вызывает электрическую поляризацию тканей верхушки: освещенная сторона получает отрицательный заряд, а затененная положительный. Эта поляризация вызывает смещение тока гормона на затененную сторону, клетки которой в результате этого растягиваются сильнее, чем освещенной, и в конечном итоге, происходит изгиб в сторону света.
Выяснено, что гормоном, вызывающим рост клеток при фототропической реакции является ИУК.
Неравномерное распределение ауксина оказывается не единственным фактором, поперечной поляризации. Возникают и другие явления, связанные с физиологией обмена веществ, в частности изменяется содержание сахара, концентрация ионов водорода в клетках освещенной стороны.
Спектр действия фототропизма определяется и качеством света. Фототропизм лучше проявляется в ультрафиолетовой области и в синей части спектра. У семенных растений фототропическая реакция вызывается коротковолновыми лучами. Фототропическая реакция зависит и от интенсивности освещения: чем слабее свет, тем дольше нужно освещать растения для получения фототропического эффекта. Произведение силы света на продолжительность его воздействия является величиной постоянной. Эта зависимость получила название закона количества раздражения. При фототропизме рецепторами являются флавопротеиды.
Фототропическая чувствительность может не проявляться, если освещать растения сразу сильным светом после их проращивания в темноте. В фототропической реакции свет выступает как раздражитель, необходимый лишь для запуска различных физиологических процессов; при этом расходуется малое количество энергии. Для ростовых процессов требуется много энергии.
Геотропизм– ориентировка осевых органов растений, которая вызвана односторонним действием силы земного тяготения. Положительный геотропизм корней вызывает его направленный рост к центру Земли, отрицательный – рост стебля от центра.
Геотропическая реакция так же, как и фототропическая, изменяется в процессе онтогенеза. Цветки некоторых растений до распускания обладают отрицательным геотропизмом, а после распускания он изменяется на положительный. Геотропизм у боковых стеблей и корней выражен меньше, чем у главных. В результате главный побег растет строго вертикально вверх, главный корень – вертикально вниз, а боковые побеги и корни располагаются под некоторым углом к ним. Это помогает растению поглощать воду и минеральные элементы из почвы, а надземным частям растений – поглощать СО2 из воздуха и избегать затенения друг друга.
Для осуществления геотропической реакции требуется определенное количество раздражения, причем неважно, будет ли это сильное раздражение в течение короткого времени или слабое в течение длительного времени. Здесь, как и в фототропизме, действует закон количества раздражителя.
Для объяснения механизма геотропизма следует обратиться к гормональной теории Холодного – Вента, согласно которой при вертикальном положении проростка ток гормонов распределяется равномерно, что и приводит к равномерному росту. При горизонтальном положении проростка гормон концентрируется на нижней стороне органа. При этом рост клетки на нижней стороне ускоряется, и стебель изгибается вверх, а у корня тормозится и он изгибается вниз. Клетки корня обладают большей чувствительностью, чем клетки стебля, поэтому одна и та же концентрация гормона на нижней стороне может подавить рост клеток корня и стимулировать растяжение клеток стебля. В результате два органа проявляют разную геотропическую реакцию.
У корней органом, воспринимающим раздражение, служит корневой чехлик. Если удалить корневой чехлик, растение утрачивает способность к геотропизму. После регенерации корневого чехлика геотропическая реакция восстанавливается.
Восприятие силы тяжести может происходить только в результате перемещения имеющих массу частиц. Такую функцию у растений могут выполнять статолиты. В 1900 г. чешский ботаник Б. Немец и немецкий физиолог Г. Геберландт независимо друг от друга создали статолитную теорию геотропизма и указали на крахмальные зерна в лейкопластах как структуры, воспринимающие силу тяжести. Такие крахмальные зерна встречаются в клетках корневого чехлика и в эндосперме – клетках первичной коры стеблей растений. При изменении положения корня крахмальные зерна в силу своей тяжести передвигаются и занимают всегда самую нижнюю часть клетки, оказывая давление на цитоплазму, которая возбуждается.
Если из лейкопластов удалить крахмал, то способность к геотропным реакциям утрачивается. Следовательно, крахмал лейкопластов играет решающую роль при выполнении ими функции статолитов.
В настоящее время предполагают, что у высших растений возможность воздействия крахмала осуществляется через давление на плазматические структуры, которые и вызывают возбуждение. Кроме крахмальных зерен аналогичную функцию, по-видимому, могут выполнять в клетках корня кристаллы белка, хлоропласты, митохондрии и другие структуры. В результате восприятия растением земного притяжения в клетках за счет статолитов наблюдается поперечная поляризация, которая проявляется в первую очередь в неравномерном распределении ауксина. Это вызывает усиление роста клеток и активное передвижение веществ.
В геотропической реакции, как и при фототропизме, различают четыре стадии: восприятие раздражения, возбуждение клетки, приводящее к накоплению гормонов, проведение возбуждения, неравномерный рост органа.
Геотропический изгиб как ростовое движение свойственен молодым частям растений; у взрослых, закончивших свой рост, он наблюдается очень редко.
Геотропическая чувствительность может меняться под влиянием факторов внешней среды. Так полегшие растения не могут подняться, если почва сильно переувлажнена и корни находятся в анаэробных условиях. При неблагоприятных условиях среды (засуха, низкая или высокая температура, действие химических веществ) крахмальные зерна исчезают, геотропическая чувствительность не проявляется.
Хемотропизм– это изгибы корней при неравномерном распределении в почве какого-либо химического вещества. Хемотропизм кроме корней свойственен пыльцевым трубкам, проросткам растений-паразитов. И здесь наблюдается положительный и отрицательный хемотропизм, который может изменяться в зависимости от концентрации и характера веществ. У корней химический сигнал воспринимает кончик корня, а изгиб находится в зоне растяжения. Механизм хемотропизма неизвестен.
Благодаря хемотропизму растение способно усваивать удобрения, избегать их избыточного накопления. Это связано с движением корня (ростом) в направлении имеющихся удобрений или при избытке их – от удобрений.
Гидротропизм –разновидность хемотропизма. При этом виде тропизма наблюдается изгибание растущих частей растений под влиянием воды. Гидрочувствительность также присуща кончику корня.
Тигмотропизм– это ростовое движение в ответ на прикосновение, давление (механическое воздействие). Типичным примером является тигмотропическое движение волосков росянки, движение усиков вьющихся растений, черешков и вершин некоторых листьев и т. д.
Термотропизм– изгиб растущего органа в направлении одностороннего действия теплового излучения.
Электротропизм – движение (изгиб), вызываемый электрическим полем, током. При этом побеги изгибаются к аноду, а корни к катоду. Изучение механизма тропизмов, в частности фототропизма и геотропизма, показало, что в них необходимо распознавать два последовательных периода : 1) возникновение разницы в физических и химических потенциалах на противоположных сторонах органа, индуцируемого односторонним светом или силой земного тяготения; 2) непосредственный тропический изгиб органа, который происходит в результате неравномерного распределения ауксинов.
Если тропизмы это ростовые движения, то настии еще и тургорные. Настии (от греческого слова нассо – уплотняю, закрываю) – движения органов растения, вызываемые (в отличие от тропизмов) раздражителями, действующими равномерно на все растения, например изменениями температуры, влажности, освещенности и т. п.
«Сонное движение» (никтинастии) некоторых цветов и листьев, когда они закрываются или открываются в ответ на изменение света (фотонастии) или температуры (термонастии) относятся к настиям потому, что внешние только запускают их, а направление зависит от внутренних факторов. Некоторые цветы, например тюльпан, закрываются ночью потому, что лепестки растут быстрее (гипонастии), а открываются в результате того, что начинает быстрее расти и верхняя часть лепестков (эпинастии).
У многих растений, особенно у бобовых (клевер) в листьях и листочках имеются особые структуры, которые называются листовыми подушечками. Быстрое изменение тургорного давления в таких клетках приводит к тому, что листовая подушечка начинает работать как шарнир, с помощью которого и происходит движение.
Гиптонастическое движение, которое происходит в ответ на прикосновение, принадлежит к самым интересным движениям (движение у мимозы). Как считают раздражение передается электрическим путем и может быть связано с движением гормонов по ксилеме.
Сейсмонастии – это быстрая реакция в ответ на сотрясение. Такой сейсмонастичной реакцией обладают листья мимозы, тычиночные нити барбариса и т. д.
Наиболее изучен механизм сейсмонастии у мимозы, у которой при сотрясении (или действии другого раздражителя) происходит заметное движение ее черешков и листьев. Опускание листьев мимозы обусловлено сокращением сочленовой подушечки, поддерживающей листовой черешок. Такого же типа подушечки, но поменьше, расположены в местах прикрепления к главному черешку четырех подчерешков, и совсем мелкие – у каждой листовой доли.
При легком встряхивании или ударе происходит опускание черешка вниз, четыре подчерешка спадают попарно назад, противоположные доли складываются вместе, и вытягиваются вверх. Опадение листа мимозы происходит вследствии сокращения в объеме сочленовой подушечки в результате потери тургора.
Протяженность периода сокращения, т. е. времени пребывания листа в движении при складывании, составляет около трех секунд. После непродолжительного покоя в опущенном состоянии лист начинает подниматься; скорость такого подъема очень невелика по сравнению со скоростью опускания; общее время так называемого периода восстановления – возвращение листа в исходное состояние – примерно 16 минут, т. е. в 300 раз больше времени сокращения. Движение определяется возникновением волны возбуждения на раздражение.
Исследователями более позднего периода было установлено, что механизм передачи раздражения у мимозы довольно сложен. Прежде всего, раздражение ударом отдельного листа чаще всего не передается другим листьям через стебель; если удар не очень сильный потенциал действия распространяется по листовому черешку лишь до сочленовой подушечки. Более слабое раздражение кончика одного из подчерешков может передаваться только соседним подчерешком. На рис. 6.22 показаны пути распространения потенциала действия при слабом раздражении одного из подчерешков.
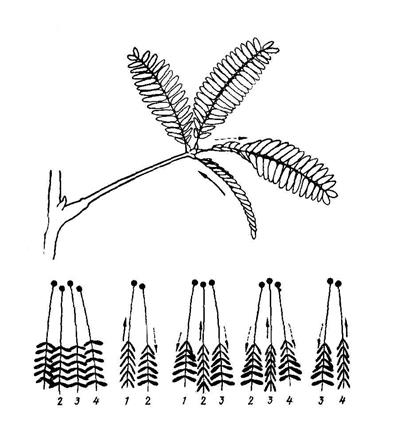
|
| Рис. 6.22. Распространение потенциала действия при раздражении одного из подчерешков листа мимозы |
Как видим, при раздражении одного из двух центральных подчерешков реагируют, в конечном счете, три подчерешка – два центральных и один крайний; если же раздражается крайний подчерешок – возбуждение передается лишь одному центральному подчерешку, соседнему с ним.
Помимо волны потенциала действия, возникающей при раздражении ударом, в проводящих пучках мимозы может распространяться и другой тип возбуждения – так называемая медленная волна (вариабельный потенциал), появляющаяся исключительно при порезах, надломах, ожогах и химических раздражениях. Природа этой волны – не электрическая; как считают, она связана с распространением так называемых раневых гормонов – специфических регуляторных веществ, возникающих в ткани при механическом повреждении.
В отличие от потенциалов действия, возникающих при ударе и, как упоминалось, обычно не распространяющихся за пределы одного листа или даже части подчерешков, медленная волна свободно минует сочленовые подушечки, омертвевшие участки стебля или черешка и т. п. Достигая стебля, медленная волна вызывает возникновение потенциала действия, распространяющегося вдоль стебля и проводящего к опусканию близлежащих листьев. В настоящее время, помимо мимозы, вариабельный потенциал обнаружен у других высших растений (тыква, виноград и др.) (рис. 6.23).
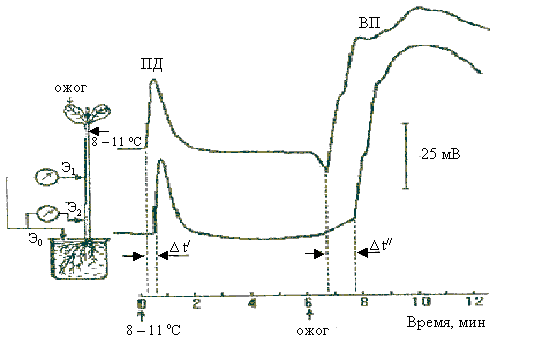
|
| Рис. 6.23. Электрические реакции проростков тыквы на охлаждение и последующий ожог: ПД – потенциал действия, ВП – вариабельный потенциал |
Иными словами, мы видим две специализированные, но взаимодействующие системы передачи возбуждения.
Заметные движения характерны и для других органов высших растений. Довольно быстро происходит движение тычинок у барбариса. В спокойном состоянии тычинки расправлены в радиальном направлении, они прилегают к лепесткам цветка. Если осторожно прикоснуться к какой-либо из них, она быстро загнется внутрь, а затем возвратиться назад (рис. 6.24, а). Движение тычинок, как показал Кабш в 1868 г. связано с электрической активностью тычинок при возбуждении. Обстоятельные исследования, выполненные впоследствии К. Умратом и Э. Бюнингом на тычиночных нитях барбариса, позволили обнаружить ПД, появляющийся при раздражении и предшествующий сгибанию тычиночной нити.
У некоторых растений способностью к движению при раздражении обладают и пестики, например у мимулуса. Рыльце его пестика снабжено лопастями, которые при раздражении закрываются. Эти движения довольно быстрые – полное соприкосновение лопастей происходит в среднем за одну минуту (рис. 6.24, б) и сопровождается возникновением ПД.

|
| Рис. 6.24. Движение тычинок и пестиков: а – цветок барбариса: 1 – пестик; 2 – тычинка до раздражения; 3 – тычинка, поднявшаяся после раздражения; б – цветок мимулуса: 1 – лопасти пестика открыты; 2 – лопасти пестика закрыты после раздражения |
Очень убедительными оказались опыты, выполненные на цветке стилидиум. Две тычинки этого цветка срастаются вместе и располагаются под некоторым углом к пестику. После раскрытия пыльников оба органа приобретают способность к движению: при механическом воздействии сдвоенная тычинка и пестик склоняются друг к другу. Электрофизиологические исследования позволили и здесь выявить электрический характер возбуждения: ПД распространяется от возбудимых клеток вдоль органа и вызывает реакцию клеток основания.
Тропизмы обнаруживаются в самом начальном этапе онтогенеза растения и проявляются в форме реакций (движений и ростовых) на действие внешних факторов еще до начала его контактов с факторами окружающей среды.
Функционирование рецепторов, принимающих участие в таких процессах как фотоморфогенез и фототропизм, создает основу для формирования хлоропластов в листе, которые разворачиваются к свету, и таким образом, и для процесса фотосинтеза.
Прикрепленная к субстрату (почве) растение выработало специфическую реакцию на одностороннее действие внешних факторов (тропизмы) или на диффузионно-действующие внешние стимулы (настии). Центры, которые участвуют в реализации таких реакций, как фототропизм или геотропизм, находятся в меристемах, а сами процессы осуществляются при участии сложного комплекса регуляторных факторов. Жизнь растения зависит от цепи экологических факторов, таких как свет, влажность, температура, сила тяжести, которые изменяются на протяжении года и неодинаковы в разных климатических зонах.
Особую группу составляют движения улавливания и захватывания, присущие насекомоядным растениям. У этих растений процессы генерации и распространения волны ПД возбуждения необходимы для обеспечения важных жизненных функций.
Известно более 500 видов насекомоядных растений, относящихся к 7 семействам. Насекомоядные растения относятся как к водным (альдрованда), так и наземным растениям (росянка, венерина мухоловка и др.). Основная функция движения органов – обеспечение недостающими элементами питания, в частности азотными соединениями.
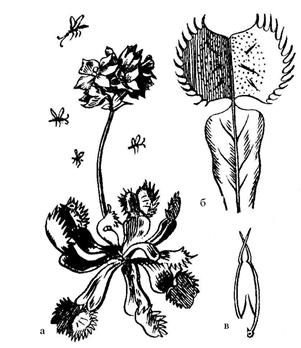
|
| Рис. 6.25. Венерина мухоловка: а – общий вид; б – раскрытый ловчий лист; в – поперечный срез листа в захлопнутом состоянии |
Например, закрытие ловчего листа венериной мухоловки происходит достаточно быстро, если между повторными раздражениями чувствительных волосков интервал составил не более 20 секунд (рис. 6.25). Почти все этапы связанные с движением и перевариванием пищи у насекомоядных растений контролируется электрическими сигналами.
Таксисы– это перемещение всей клетки или всего организма, которое вызывается и направляется определенными внешними стимулами. Как и тропизмы таксисы также делятся на положительные и отрицательные; кроме того, их можно классифицировать в соответствии с природой раздражителя: сила тяжести, света, химические воздействия (тигмо-, фото-, хемотаксисы).
Многие из физиологических реакций (движений) осуществляются потому, что в растениях имеются специфические структуры – рецепторы (лучше биологические мишени), воспринимающие сигналы. К числу таких рецепторов, прежде всего, относится комплекс каротиноидов и флавинов, которые осуществляют ориентацию стебля в пространстве. Крахмальные зерна (статолиты) участвуют в регулировании геотропизма корня. Интересно, что эти вещества близки по строению к рецепторам высших животных и человека. Каротиноиды подобны витамину А, хлорофилл имеет сходство с гемом эритроцитов. Рецепторы чувствительно реагируют на смену внешних факторов и передают сигнал об этой смене растительному организму. При этом часто происходит изменение структуры самого рецептора.
Дата добавления: 2016-07-11; просмотров: 4785;
Поиск по сайту
Узнать еще
- I.6.2 ГИДРОАЭРОСТАТИКА
- III. Экстраполяция по темпу роста.
- XX съезд КПСС и осуждение культа личности. Экономические реформы конца 50-х – начала 60-х гг. Причины их неудач. Замедление экономического роста.
- А. Электростатическое экранирование
- Адаптация растений и животных к световому режиму.
- Алгоритмы группы KWE
- Алгоритмы диагностики
- Алгоритмы динамического управления памятью

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
