Зависимость роста растений от экзогенных факторов
Рост, являясь интегральной функцией растения, зависит от влияния абиотических факторов внешней среды: света (его интенсивности, спектрального состава, продолжительности и периодичности), температуры (величины и периодичности), питательных веществ (количества и сбалансированности), механических воздействий ветер и др.).
Регуляция роста светом. Разносторонние процессы, которые определяют влияние света на форму растений, были названы фотоморфогенезом.
Растения, выросшие в темноте, называются этиолированными. Они отличаются от растений, выращиваемых на свету, рядом морфологических и биохимических особенностей.
Программа развития нормального зеленого растения контролируется специальными световыми реакциями, лежащими в основе фотоморфогенеза. Хлорофилл и разные вспомогательные пигменты фотосинтетического аппарата не играют в этих процессах главной роли, посколько начальные этапы развития растений на свету не могут осуществляться за счет фотосинтеза: структурная и биохимическая основы фотосинтетического аппарата в этот период еще не сформированы. Для устранения эффекта этиоляции и включения процессов фотоморфогенеза достаточно ежедневного 5–10 минутного освещения растений.
Термин «фотоморфогенез» объединяет процессы, которые не зависят от направления и периодического освещения, а определяются длиной волны и интенсивностью светового потока. Вероятно, в процессах фотоморфогенеза растений играют роль определенные соединения – пигменты.
Содержание этих пигментов в растениях очень малое, поэтому количество энергии, необходимое для насыщения соответствующих фотопроцессов, на несколько порядков ниже, чем при фотосинтезе. В то время как при фотосинтезе 8–10 квантов света необходимо для выделения только одной молекулы О2, такое же количество квантов на одну клетку может полностью определить репродуктивную судьбу растения или направление роста всего стебля.
Реакция растений на длительность светового периода (или длину дня) разная. Эта реакция растений на продолжительность дня получила название фотопериодизма. Выделяют растения короткого дня, когда растения зацветают при небольшой продолжительности освещения – менее некоторой критической величины. Когда же длина дня превышает определенную критическую величину – это растения длинного дня. Имеется и группа растений, цветение которых не зависит от длины дня – нейтральные растения.
Критический фотопериод у разных видов и даже сортов растений как короткого, так и длинного дня значительно варьирует. Эти различия в критическом фотопериодизме играют важную роль в размещении растений на Земле.
После открытия фотопериодизма выявили, что многие растения реагируют на длительность непрерывной темноты, а не на протяженность светового периода. Иными словами, так называемые растения короткого дня – это на самом деле растения длинной ночи. Для закладки цветковых бугорков им необходимо определенная минимальная длительность темноты, которая не прерывается светом. Подобным образом растение длинного дня может быть в действительности растением короткой ночи: оно будет цвести только в том случае, если ночной период не продолжительней некоторого максимума.
Всего только один подходящий период темноты может привести растение короткого дня к цветению, даже тогда, когда последующие темновые периоды недостаточно длительны. Этот феномен известен как фотопериодическая индукция. У многих растений длинного дня имеет место подобный, но обратно направленный феномен: прерывание очень длительного темнового периода вспышкой света приводит к индукции и заложению цветков.
Последнее говорит о том, что растения длинного дня и короткого дня, вероятно, владеют одинаковыми фотопериодическим механизмом, но последний каким-то образом действует в разных направлениях.
Это нашло дальнейшее подтверждение при изучении действия света разной длины волны – спектр действия. Для ингибирования цветения короткодневных растений и для активации цветения длиннодневных растений во всех случаях был определен максимум активности в красной области спектра (около 660 нм) при почти полной неэффективности других длин волн. Подобие спектров позволило считать, что зацветание растений как короткого, так и длинного дня контролируется одним и тем же пигментом.
Далее, свет сильно влияет на прорастание семян. Некоторые семена плохо прорастают в полной темноте, но быстро и хорошо при экспонировании на свету на протяжении нескольких минут. Оказалось, что спектр действия для этого эффекта подобен спектру действия для цветения.
Проросток (гороха), который выращивается в полной темноте, имеет очень длинный тонкий стебель, апикальный изгиб и почти не распустившиеся листья. Когда такой этиолированный проросток выставить только на короткое время на свет, листья после этого разворачиваются, изгиб начинает выпрямляться, удлинение стебля замедляется. Поэтому, появилась возможность заключить, что такие разносторонние реакции как прорастание семян, изменение этиолированного проростка (его деэтиолирование) и индукция цветения регулируются одним и тем же пигментом – рецептором.
Как определили, таким пигментом является фитохром. Фитохром – от греческого слова «растение» и «краситель». Фитохром представлен растворимыми в воде соединениями, относящимися к билихромпротеинам. Как и у фикобилинпротеинов (фикоцианобелин), хроматофор (пигментная часть молекулы) фитохрома представлен незамкнутыми порфиринами (тетрапирол). Другая часть молекулы образуется бесцветным белком. После поглощения фитохромом света с определенной длинной волны форма хроматофора изменяется, и, это в свою очередь, изменяет форму белкового компонента. Молекулярная масса фитохрома колеблется в зависимости от вида растения от 120 до 170 кДа. В составе фитохрома – одна полипептидная цепь. Существует, как предполагают, фитохром в клетке в виде димера.
Фитохром регулирует многие физиологические реакции, наблюдаемые при поглощении малых доз красного и дальнего красного света растениями, которые находились в полной темноте.
Фитохром находится в растениях в двух формах: Р660 – поглощает красные лучи и превращается в Р730; последний поглощает дальние красные лучи и переходит в Р660 (рис. 6.16). Р660 (Рк) – физиологически неактивный.
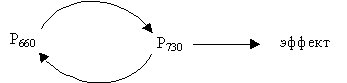
|
| Рис. 6.16. Схема биологического ответа при действии красного и дальнего красного света |
Структура фитохрома представлена на рис. 6.17.

|
| Рис. 6.17. Структура фитохрома |
Реакции, управляемые фитохромом, зависят от концентрации Р730 (Рдк).
Полагают, что Ф660 содержит на один протон больше, чем Ф730 (см. рис. 6.17). Эта разница влияет на конформацию хромотофора и белковой молекулы, с которой он связан.
Фитохром присутствует в растениях, как уже отмечалось, в небольших количествах: его концентрация составляет около 1 мкМ. При этом он размещен в растении неравномерно, больше всего его в растущих, делящихся тканях, например в клетках меристемы. В клетках стебля он практически отсутствует.
До освещения красным светом фитохром обычно распределен по всей цитоплазме и ее мембранах, а после короткого освещения выявляется только в определенных частях клетки. Вероятно, эти части – ни ядра, ни пластиды, ни митохондрии, а, скорее, ЭР, разбросанный по всей цитоплазме. Длительное воздействие света приводит к появлению фитохрома уже и в ядрах. Подтверждают эти выводы эксперименты с изолированными субклеточными фракциями; однако, не все согласны с интерпретацией полученных данных.
Все же данные физиологических опытов позволяют предполагать, что фитохром может находиться в клетке во многих местах в соответствии с его многочисленными функциями. Вероятно, он локализован внутри этиопластов и митохондрий, или в их наружных мембранах, так как изолированные органеллы реагируют на воздействие красным и дальним красным светом, после их выделения. Предполагают, что фитохром находится (или каким-то образом связан) в плазмалемме.
Считают даже, что хроматофору фитохрома свойственна определенная ориентация в плазмалемме. Кроме того, вероятно, что фитохром изменяет свою ориентацию в мембране на 90 о при фототрансформации
Обычно реакции достигают насыщения, когда 50 % фитохрома представлена формой Р730, и продолжается до той поры, пока имеется достаточное количество Р730.
Превращение Р660 ↔ Р730 происходит также и in vitro. Как считают, оно связано с конформационными изменениями участков белковой части молекулы фитохрома. Этот процесс происходит через промежуточные этапы, потому что пигмент исчезает быстрее, чем появляется другой.
Из-за того, что Р730 неустойчив, его количество в темноте уменьшается, что объясняется медленным тепловым превращением в Р660 и довольно быстрым, по всей вероятности, окислительным разрушением, катализируемым металлами. Очень маленькое количество Р730 остается при выдерживании растений в темноте.
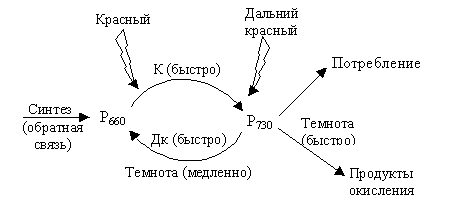
|
| Рис. 6.18. Схема превращения фитохрома |
Потери фитохрома компенсируются in vivo его синтезом, который начинается по принципу обратной связи, когда суммарное количество фитохромов (Р660 + Р730) становится ниже порогового (рис. 6.18). Синтез пигмента (незамкнутого порфирина) происходит, как и синтез хлорофилла (циклического порфирина), через δ-аминолевулиновую кислоту.
Под влиянием Дк света Р730 быстро и почти целиком (около 90 %) превращается в Р660, а при красном свете Р660 превращается в Р730 примерно на 80 %.
Солнечный свет содержит приблизительно одинаковые доли К и Дк. Это приводит к равновесию Р730/Р660, при котором содержание Р730 доходит до 50 % (во всяком случае, больше, чем в темноте). Поэтому в реакциях, регулируемых фитохромом, солнечный свет действует как красный.
Важная роль фитохрома обусловлена тем, что это единственный из растительных пигментов, который использует световые сигналы для управления развитием растения. Ряд ферментативных реакций, связанных с процессом фотосинтеза регулируются фитохромом.
В соответствии с общими представлениями, впервые высказанными Х. Мором (1966 г.), действие фитохрома связано в первую очередь с регуляторным влиянием Дк-формы на активность так называемых фотоиндуцибильных генов. В общем виде генетический путь трансдукции может быть представлен следующей схемой:
К → Дк → активация или репрессии гена → рост или снижение содержания РНК → модификация содержания белкового продукта → биологический эффект.
Кроме того, на основании полученных данных считают, что Са2+ и кальмодулин принимают участие в передаче сигнала от Дк-формы фитохрома к генетическому аппарату, а так же в этом процессе участвует и G-белок. Генетический механизм обеспечивает у растений медленные фитохромные ответы.
Однако, быстрые фитохроминдуцируемые эффекты (фотоиндуцируемые изменения потенциала, набухание хлоропластов и т. д.), вероятно связаны с модификацией мембранных структур. В механизмах мембранной трансдукции фитохромного сигнала важную роль играет Са2+.
Мембранный механизм передачи фитохромного действия имеет следующий вид:
К → Дк → изменение проницаемости мембраны для ионов Са2+ → временное изменение концентрации Са2+ в цитоплазме → включение кальций-зависимой биохимической системы → биологический ответ.
Кальциевый поток действует на активность кальмодулина, последний изменяет активность ферментов, например, протеинкиназ. Через действие катаболических ферментов может происходить активация фитогормонов, запускающих механизм ростовой фотореакции.
Так состояние фитохрома может регулировать качественный и количественный состав гормонов четырех типов – этилена, цитокининов, ауксина и гиббереллинов – в разных органах растения. Кроме того, длительное облучение иногда контролирует уровень АБК. В некоторых случаях внесение гормона может снижать эффект трансформации фитохрома (например действие гиббереллина при деэтиляции).
В природе фитохром, вероятно, контролирует форму растений и прорастание семян под пологом листвы, так как листья очень сильно поглощают свет в области 660 нм и относительно слабо – 730 нм. Фитохром может регулировать прорастание семян, деэтилирование, цветение, опадение листьев и даже старение.
Синий свет также регулирует фотоморфогенетические реакции, в частности движение устьиц, фототропизм, ранние этапы синтеза хлорофилла и каротиноидов, движение листьев за солнцем, фототаксис хлоропластов, угнетение роста гипокотилей и стеблей в длину. Фоторецепторы, участвующие в восприятии синего света, называют криптохромами.
У растений арабидопсиса выявлены соединения (криптохром 1, криптохром 2 и фототропин), которые воспринимают и передают сигналы синего света. Криптохропы – рецепторы синего света, локализованные в ядре и цитоплазме, включающие белковую и хромофорную составляющие. Функции хромофоров у криптохромов выполняют флавины (ФАД, деазафлавин) и (или) птерин.
Криптохромная система замедляет рост гипокотеля (или междоузлия) на свету, помогает фитохрому контролировать процессы диэтиоляции. Обе фоторецепторные системы (криптохром и фитохром), действующие в ядерном компартменте, дополняют друг друга, что сказывается в интеграции информации о физиологически важных областях спектра – синей и красной.
Фототропин представляет собой ассоциированный с мембраной белок, т. е. является мебранным рецептором синего света, функцию хромофора у которого выполняет флавин ФМН. Фототропин ответственен за фототропизм, хемотропизм и потенциалзависимые мембранные процессы.
Характер ответных реакций растений при освещении синим светом определяется его интенсивностью и длиной волны.
У папоротника Adiantum capillus-veneris был найден ген, гомологичный как фитохромам, так и фототропинам. «Двойной» фоторецептор был назван суперхромом. Как и фототропин, суперхром обладает протеинкиназной активностью.
Следовательно, если у одних растений фототропин взаимодействует с фитохромами опосредованно, то у Adiantum проблема взаимодействия красного и синего световых сигналов решена на уровне «двойного» рецептора.
Таким образом, конечная форма зеленого растения, обусловленная в целом его генами, определяется участием света, который действует через разные фоторецепторы.
Влияние температуры. Яровизация. Стратификация.Реакции на температуру, так же как на свет, у растений могут быть количественными и качественными. Скорость почти всех химических процессов с повышением температуры возрастает, достигает некоторого оптимума, а затем снижается. В отличие от этого многие процессы онтогенеза, например, прорастание семян, прерывание покоя почек, часто регулируется по закону «все или ничего». Это означает, что, например для низкотемпературной индукции требуется воздействие на протяжении какого-то минимального периода; приведенная закономерность напоминает фотопериодическую индукцию, при которой также необходимы определенные периоды темноты.
Влияние температуры, как мы уже отмечали, на физиолого-биохимические процессы характеризуют температурным коэффициентом:
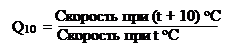 |
При анализе зависимости роста растений от температуры выделяют три кардинальные точки: минимум (рост только начинается), оптимум (наиболее благоприятный период для роста) и максимум (прекращение роста). Кардинальные точки для различных растений неодинаковы. У теплолюбивых растений (кукуруза, огурец, дыня, тыква), все точки смещены в сторону более высоких температур.
В среднем минимальные температуры для роста растений умеренной зоны –5 – +15, оптимальные – +25 – +35, максимальные – +37 – +44 0С. Растения подразделяются на теплолюбивые и холодостойкие.
Кривые зависимости скорости роста растения от температуры представлены на рис. 6.19.
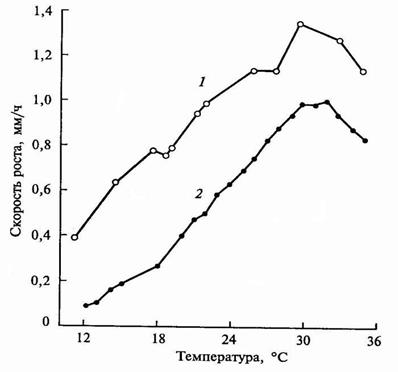
|
| Рис. 6.19. Влияние температуры на рост корней гороха (1) и колеоптилей кукурузы (2) |
Пока еще окончательно неясно почему большинство растений повреждается температурами выше ~ 30 оС, тогда как ферменты или органеллы, выделенные из растений, при таких температурах обычно не повреждаются. Одно из возможных объяснений заключается в том, что мембраны клеток или органелл чувствительны к изменениям температуры из-за плавкости или затвердения жирных кислот в фосфолипидах. Известно, что растения при низких температурах синтезируют больше ненасыщенных жиров с более низкой температурой плавления. Обратное явление наблюдается при высоких температурах. Другая возможность состоит в том, что при повышенных температурах какие-то материалы, необходимые для роста, очень быстро разрушаются или не образуются в необходимых количествах. Во многих организмах имеются даже гены, «чувствительные к температуре».
У плесневого гриба Neurospora ген, ответственный за образование витамина В2 (рибофлавин), хорошо функционирует при низких температурах, но плохо действует при более высоких. Поэтому при 35 оС гриб должен получать рибофлавин из вне. Подобным же образом может происходить у высших растений при недостатке витаминов, аминокислот, гормонов и т. д. Таким образом, можно было бы улучшить рост путем внесения необходимых веществ.
Очень маленькая скорость химических реакций в растении при низких температурах обеспечивает координацию изменения роста с климатическими изменениями. Кроме того, температура влияет на многие процессы, чувствительные к фотосинтезу. Некоторые растения можно заставить цвести с помощью длинных темных ночей или низких ночных температур.
Низкая температура может вызвать прорастание семян, сдвиги температуры могут прервать покой почек и подготовку к закладке цветочных бугорков.
Для появления всходов требуются более высокие температуры, чем для прорастания семян (табл. 6.2).
Таблица 6.2
Дата добавления: 2016-07-11; просмотров: 3473;
Поиск по сайту
Узнать еще
- I.6.2 ГИДРОАЭРОСТАТИКА
- III. Экстраполяция по темпу роста.
- Independence of the Seas. (Независимость морей)
- Pиc. 67. Зависимость скорости осаждения от величины катодно-анодного отношения
- XX съезд КПСС и осуждение культа личности. Экономические реформы конца 50-х – начала 60-х гг. Причины их неудач. Замедление экономического роста.
- А) Зависимость удельной тяги и удельного расхода топлива ТРДФ от параметров рабочего процесса
- А. Электростатическое экранирование
- Автоматическая работа и взаимозависимость клеток организма

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
