Гомеостаз и поведение
Понятие физиологической стабильности неотделимо от концепции Клода Бернара (Claude Bernard. 1859) о внутренней среде. Он установил, что уровень сахара в крови остается постоянным независимо от того, голодало животное, только что получало мясо или потребляло корм, содержащий сахар. Он постулировал наличие некоторого процесса регуляции и контроля для поддержания постоянства внутренней среды. Он понимал также, что животное, способное регулировать свою внутреннюю среду при колебаниях внешней среды, способно использовать большее разнообразие потенциальных местообитаний.
Животных можно приблизительно разделить на конформеров, допускающих влияние на свою внутреннюю среду внешних факторов, и на регуляторов, которые удерживают ее в состоянии, в значительной степени независимом от внешних условий. Процессы, посредством которых регуляторы управляют своим внутренним состоянием, объединяются термином гомеостаз.
Гомеостаз
Этот термин впервые был применен американским физиологом Кэнноном (Cannon. 1932), который писал: «Координированные физиологические процессы, которые поддерживают большую часть устойчивых состояний в организме, так сложны и настолько специфичны для живых существ, поскольку могут включать совместную работу головного мозга и нервов, сердца, легких, почек и селезенки, - что я предложил специальное название для этих состояний, а именно гомеостаз». Кэннон рассмотрел ситуацию, в которой сенсорные процессы, следящие за внутренним состоянием организма, вызывают соответствующее действие, как только внутреннее состояние отклонялось от заданного или оптимального. Так, например, когда температура человеческого тела поднимается выше 37 С, вступают в действие такие охлаждающие механизмы, как прилив крови к коже и потоотделение. Когда температура падает ниже оптимального уровня, включаются согревающие механизмы, например дрожь. Пользуясь множеством таких тонко настроенных механизмов, человек достигает точной терморегуляции и теплового гомеостаза.
Хотя типы регуляторных механизмов, рассмотренные Кэнноном, как теперь известно, широко распространены в животном мире и включают самые разнообразные физиологические процессы, в регуляции внутренней среды участвуют не только они. Так, например, до недавнего времени считали, что животные усиленно пьют при высокой окружающей температуре в результате обезвоживания, которое возникает при таких охлаждающих реакциях, как потоотделение и одышка. Эта точка зрения полностью соответствует изложенной выше теории гомеостаза: испарение воды необходимо для поддержания теплового гомеостаза в жаркой среде, а это нарушает в организме водный баланс, восстановление которого требует усиленного питья. Однако теперь мы знаем, что у таких видов, как крыса и голубь, питье прямая реакция на температурное изменение, опережающая любое нарушение водного баланса в результате терморегуляции, а не ответ на нарушение.
как считает регуляторная теория (Budgell, 1970а, b). Иными словами, животные пьют, чтобы запастись водой для терморегуляции.
Роль поведения в регуляции внутренней среды значительно отличается у разных видов и зависит от обстоятельств. Питье, например, нужно для поддержания гомеостаза у многих видов, так как физиологические механизмы не могут защитить их от гибели в результате обезвоживания после длительного лишения воды. Но некоторые виды, например монгольская песчанка (Meriones unguiculatus) и волнистый попугайчик (Melopsittacus undulatus) (Cade, Dybas, 1962), выживают без воды неограниченное время. Благодаря большой эффективности своих сохраняющих воду механизмов они могут жить, пользуясь влагой, содержащейся в поедаемых семенах. Другие виды, например водные, не нуждаются в особом поведении для получения воды, необходимой для поддержания гомеостаза.
Во многих случаях роль поведения в поддержании гомеостаза в норме ничтожна. Однако опыты с хирургическим вмешательством, нарушающим нормальные физиологические механизмы гомеостаза, показывают, что животные часто способны к соответствующему поведению, даже если не пользуются им в обычной жизни. Так, в работе Рихтера (Richter, 1942-1943) показано, что, если нарушить тепловой гомеостаз у крыс, удалив у них щитовидную железу, животные реагируют сооружением более теплых гнезд и другими формами поведенческой терморегуляции, возможными в условиях эксперимента. Точно так же удаление надпочечников, участвующих в поддержании солевого баланса, заставляет крыс предпочитать более соленую пищу и воду.
В других случаях участие в гомеостатических реакциях специальных органов необязательно. Так, крысы, содержащиеся на диете с недостатком витаминов, способны выбирать пищу с нужным их содержанием, хотя и не могут узнавать по вкусу о наличии витаминов в корме. Как будет показано в гл. 18, они способны научиться распознавать пищу, при которой чувствуют себя лучше (Revusky, Garcia,
1970; Rozin, Kalat, 1971). Ясно, что механизмов поддержания внутренней стабильности много и они разные, поэтому неправильно думать, что гомеостаз обязательно предполагает простой регуляторный процесс типа обратной связи. Кроме того, неверно считать, что гомеостаз означает просто постоянство внутренней среды. Между конформерами и регуляторами существует много промежуточных форм, причем вид может быть способен регулировать одну функцию организма и не способен регулировать другие. Это хорошо видно на примере терморегуляции у животных.
Терморегуляция
Для большинства животных существует оптимальная температура тела, при которой они функционируют наиболее эффективно. Ниже этой температуры их метаболизм постепенно замедляется, мышечная активность уменьшается и животное становится вялым. Выше оптимальной температуры метаболизм быстро усиливается, и для сохранения его нужного уровня могут потребоваться слишком большие затраты. Кроме того, существует верхний температурный предел жизнеспособности организма. Для большинства видов этот предел, по-видимому, близок к 47°С.
Большинство животных способно до некоторой степени влиять на температуру тела с помощью специальных физиологических механизмов или соответствующего поведения. В обоих случаях животному необходимо определять внешнюю температуру, температуру своего тела или обе сразу при помощи различных сенсорных процессов, составляющих так называемую терморецепцию (см. гл. 12).
Метаболические реакции организма непрерывно производят тепло, и чем активнее животное, тем больше образуется у него тепла. Однако иногда в холодной внешней среде калорий, получаемых от нормального метаболизма и активности, недостаточно для противодействия холоду, и тогда животное может производить дополнительное тепло, усиливая метаболизм, и принимать меры, снижающие по-
тери тепла телом. Многие беспозвоночные холоднокровны в том смысле, что температура их тела соответствует температуре окружающей среды. Поскольку интенсивность метаболических реакции определяется температурой, при которой они протекают, такие животные вынуждены снижать активность, когда температура тела падает. Однако у некоторых беспозвоночных, например у мокрицы Porcellio scaber и многоножек (Myriapoda). падение температуры стимулирует дополнительную активность, и таким образом они способны поддерживать более высокую температуру тела, чем температура среды. У теплокровных животных образование калорий можно интенсифицировать, усилив мышечную активность, например дрожью, и прямым воздействием тиреоидных гормонов на интенсивность метаболизма. Прием пищи тоже может усиливать образование тепла, так как оно выделяется при пищеварении. В холодных условиях многие животные усиленно потребляют пищу.
Животных, получающих тепло главным образом из внешних источников, например от солнца, обычно называют экзотермными, а использующих преимущественно энергию внутренних процессов, - эндотермными. Экзотермные животные иногда усиливают свое нагревание солнечными лучами, меняя окраску, поскольку темные объекты поглощают больше лучистой энергии, чем светлые. Некоторые виды ящериц способны менять окраску в соответствии со своими тепловыми потребностями. Например, пустынная игуана Dipsosaurus dorsalis имеет темную окраску ранним утром, когда температура тела низкая, но постепенно бледнеет по мере повышения его температуры, достигая средней стадии изменения окраски приблизительно при 40°С. Подобным же образом некоторые черепахи, чтобы получить больше тепла, выставляют на солнце свои черные лапы.
Метаболические реакции непрерывно генерируют тепло, и животные легко перегреваются, особенно когда они активны. Перегревание также легко наступает в особо жаркой среде или когда нарушено отведение тепла. Поскольку летальная температура тела у многих животных не намного выше нормальной, охлаждающие механизмы должны быть особенно быстрыми и эффективными.
Тело теряет тепло четырьмя главными способами: проводимостью, конвекцией, излучением и испарением. Проводимость, или теплопроводность, - это перенос тепла между областями с различной температурой в твердых и жидких телах. Она может иметь место в тканях организма или между ними и внешним объектом, например почвой. Проводимость может быть снижена теплоизоляцией, создаваемой слоями жира внутри тела, и воздухом, удерживаемым волосяным или перьевым покровом на поверхности тела. Конвекция - это перенос тепла в текучей среде. Тепло теряется вследствие притока теплой крови из внутренней части тела к более холодным поверхностным тканям. Таким образом, направление кровотока на периферию служит важным средством терморегуляции. Излучение- это форма передачи тепла, не зависящая от теплоносителя и происходящая даже в вакууме. Потеря тепла при излучении приблизительно пропорциональна разнице температур между животным и средой. За счет излучения тепло и приобретается, и теряется, причем окраска животного мало влияет на потерю тепла, но, как было сказано, важна для его приобретения. Так, черные животные поглощают больше солнечного тепла, чем белые, которые сильнее отражают свет. Испарениеводы с влажных поверхностей тела животного связано с затратой тепла и у многих видов служит важным способом охлаждения.
Разные виды способны регулировать количество отдаваемого тепла этими четырьмя способами в разной степени. Отчасти они регулируют потери тепла за счет проводимости, меняя свою теплоизоляцию. Долговременная регуляция может осуществляться усилением отложения жира и роста шерсти зимой и замедлением этих процессов летом. Кратковременных изменений животные достигают, взъерошивая или укладывая шерсть и перья (рис. 15.1), чем регулируется количество удерживаемого в них воздуха, а также меняя положение тела в зависи-
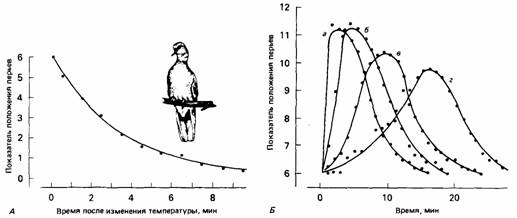
|
| Рис. 15.1. Движение перьев у египетской горлицы (Streptopelia risoria) при внезапном повышении окружающей температуры (А) и после выпивания 10 см3 холодной воды (Б). Учитывается среднее положение перьев по всей поверхности тела (см. рис. 22.10). Кривые а, 6, в, г - реакции на питье с разной скоростью. (По McFarland, Budgell, 1970.) |
мости от погоды. На поверхностную теплоизоляцию влияют ветер и влажность. Влажная шерсть повышает теплопотери за счет проводимости. Когда млекопитающее лежит или сидит на земле, шерсть уплотняется и содержит меньше воздуха. Тепло в этом случае уходит в почву, в особенности если она влажная. Часто можно видеть, как перед дождем коровы опускаются на землю для отдыха, сохраняя при этом тепло, которое было бы потерянным через некоторое время - когда земля станет влажной.
Некоторые животные могут усиливать излучение тепла с помощью поведенческих средств. Так, некоторые манящие крабы (Uca), суслики (Citellus) и другие роющие животные в жаркую погоду периодически прячутся в своих прохладных норах, где перегретое тело может охлаждаться путем теплоизлучения. Эффективную поверхность тела можно уменьшить, свернувшись клубком или сбившись в кучу с другими особями того же вида, что снижает потери тепла за счет излучения.
Испарение воды через кожу не контролируется у амфибий, рептилий и птиц; у млекопитающих оно регулируется потовыми железами, которыми обладают все высшие млекопитающие, кроме грызунов и зайцеобразных (кроликов). У человека функционирование потовых желез на ладонях находится под контролем эмоций, а на остальной поверхности тела они в норме реагируют на терморегуляцию. Потоотделение контролируется терморецепторами в головном мозгу, а не в коже. Так, люди обычно потеют при физической работе, но не обязательно, когда сидят у жаркого огня. Крысы и некоторые другие животные усиливают охлаждающее испарение, увлажняя поверхность тела слюной или смачивая себя водой, как это делают слоны.
Испарение через дыхание у большинства животных до некоторой степени регулируется. Крокодилы, змеи и некоторые ящерицы в жару широко раскрывают пасть. Пустынная игуана Dipsosaurus учащает дыхание подобно собаке. Из-за того что животные при респираторном испарении теряют очень много воды, они прибегают к нему только в крайних случаях. Птицы и млекопитающие учащают дыхание, только когда температура тела приближается к летальному уровню. Птицы в полете генерируют много тепла и для рассеяния его используют главным образом респираторное испарение. У верблюда не бывает тепловой одышки, но он охлаждается, излучая тепло ночью. Верблюды запасают не так много воды, чтобы позволить себе тратить ее для охлаждения (рис. 15.2).
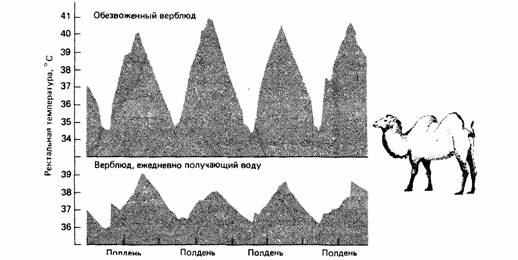
|
| Рис. 15.2. При обезвоживании температура тела у верблюдов днем повышается, так что им не приходится тратить воду на терморегуляцию. Холодной пустынной ночью верблюд теряет избыток тепла, накопленный днем. (По Schmidt-Nielsen. 1964) |
Экзотермные животные способны регулировать температуру тела только в ограниченной степени. У амфибий тело остается прохладным благодаря испарению воды с его поверхности. Леопардовая лягушка (Rana pipiens) может сохранять температуру тела 36,8° С при температуре окружающей среды 50°С. У пресмыкающихся способность к самоохлаждению более ограничена, и они стремятся избегать очень жарких мест. Намибская песчаная ящерица (Aporosaura anchietae) зарывается в песок, когда полуденная температура превышает 40°C (рис. 15.3).
Истинным тепловым гомеостазом обладают птицы н млекопитающие, которые являются эндотермными. способными сохранять постоянную температуру тела, несмотря на флуктуации окружающей температуры. Высокая интенсивность их метаболизма создает внутренний источник тепла, а покровы препятствуют его неконтролируемому рассеянию. У птиц и млекопитающих температура тела обычно выше окружающей. Головной мозг получает информацию о температуре тела и может контролировав механизмы согревания и охлаждения. Когда температура мозга становится слишком высокой, активируются механизмы охлаждения, а если она падает слишком низко, теплопотери уменьшаются и вступают в действие механизмы согревания. Этот принцип обратной связи такой же, как у термостатически регулируемого электрического нагревателя (рис. 15.4).
Тонкий контроль температуры тела происходит у человека с его системой раннего предупреждения, состоящей из множества терморецепторов в коже. На основе получаемой от них информации людиспособны принимать предварительные меры, не допуская излишних колебаний температуры тела. Контролируемые изменения температуры у птиц и млекопитающих все же происходят, часто в зависимости от времени суток. У человека средняя температура тела равна 36.7° С рано утром и 37.5° C в конце дня. Многие эндотермные животные допускают некоторые колебания внутренней температуры, вероятно, ради сохранения энергии.
Водный баланс
Все животные нуждаются в воде для поддержания метаболических процессов. Они постоянно теряют воду множеством разных способов, включая экскрецию и
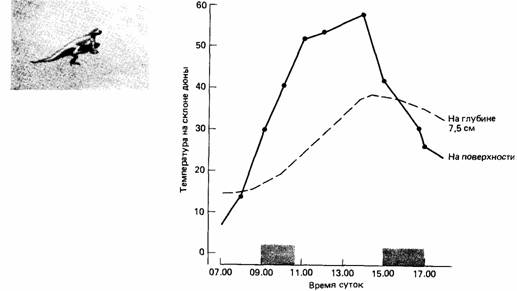
|
| Рис. 15.3. Ящерица Aporosaura живет на песчаных дюнах, на поверхности которых температура заметно колеблется. При слишком низкой или слишком высокой температуре ящерица зарывается в песок. Обычно это означает один период активности на поверхности утром и еще один - после полудня. Активные периоды обозначены серыми прямоугольниками. (Louw, Holm, 1972; фотография Gideon Louw.) |
испарение с поверхности тела. Вода в теле безпозвоночных распределена между различными компартментами (рис. 15.5). Потери происходят из кровеносных сосудов. Например, в легких вода теряется за счет испарения. Они пронизаны густой сетью капилляров, а газообмен происходит через тонкие мембраны, смоченные плазмой, в которой растворяются газы. Выдыхаемый воздух почти всегда содержит воды больше, чем вдыхаемый, и эта вода поступает из сосудистого русла. Как уже говорилось, у многих животных испарение при дыхании - существенный способ охлаждения; в частности, для птиц оно очень важно во время полета. Терморегуляция, при которой происходит потоотделение или смачивание тела слюной, тоже требует расхода воды.
Почки фильтруют плазму крови, удаляют из нее ненужные вещества и выделяют их с мочой. Следовательно, потеря некоторого количества воды при экскреции неизбежна, но, как мы увидим, здесь возможна также известная экономия. Вода теряется также во время дефекации, но при дефиците воды многие животные реабсорбируют воду из тонкого кишечника и выделяют очень сухой кал. Потеря воды из кровеносных сосудов приводит ее к некоторому перераспределению между всеми компартментами тела, но в конечном счете недостающая вода должна быть возмещена.
Большинство животных получают воду во время питья, но амфибии и некоторые насекомые поглощают ее кожей. Ряд животных, особенно редко встречающих воду, способны выпивать ее в огромных количествах. В то время как человек при сильной жажде может выпить один литр воды за минуту и, возможно, три литра за десять минут, один верблюд-самец массой 352 кг выпил 104 л за один прием. Попав в рот, вода затем проходит через пищевод в желудок и дальше в кишечник. Отсюда она может попасгь в кровь посредством осмоса. Если концентрация солей в жидкости кишечника ниже, чем в крови, вода проникнет в кровь. Но если эта концентрация выше (например, после приема соленой пищи или воды), вода
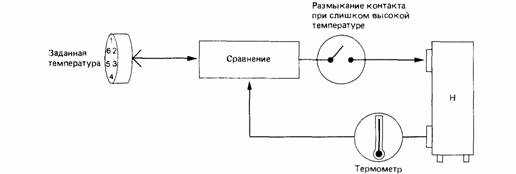
|
| Рис. 15.4. Принцип обратной связи, действующий в простом термостатическом электpонагревателе. Когда температура достигает заданного уровня, контакт прерывается, нагреватель выключается и температура падает. Koгда она упадет ниже заданного уровня, нагреватель снова включится. |
может мигрировать в обратном направлении и в сосудах наступит временное обезвоживание.
Выход воды из кровеносных сосудов приводит к изменению концентрации солей во внеклеточном компартменте, что вызывает некоторое перераспределение воды между ним и клетками; в результате клетки частично обезвоживаются и сморщиваются. Эти изменения обнаруживаются специальными клетками в головном мозгу, называемыми осморецепторами. Чувствительны ли осморецепторы к изменениям во внеклеточной жидкости или реагируют на свое собственное сморщивание при дегидратации, пока не ясно (см. обзор этого вопроса - Toates, 1980). Известно, что они находятся в гипоталамической области мозга, и их стимуляция оказывает два главных действия: усиленный поиск воды для питья и активацию разных механизмов ее сохранения.
В гипоталамусе находятся осморецепторы. которые контролируют выделение антидиуретического гормона из расположенного непосредственно под ним гипофиза (рис. 15.6). Присутствие антидиуретического гормона в кровотоке ведет к понижению количества и повышению концентрации мочи, выделяемой почками. Повреждение гипофиза или связанных с ним областей гипоталамуса вызывает не-
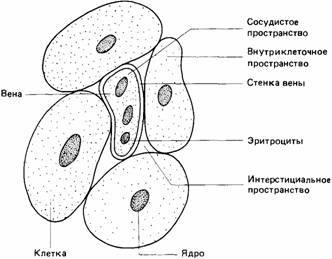
| Рис. 15.5. Распределение воды между внутриклеточным и внеклеточным компартментами тела. Внеклеточный компартмент включает сосудистое иинтерстициальное пространства. |
| Рис. 15.6. Схема строения гипофиза человека. Гипоталамус (Г) снабжается кровью из внутренних сонных артерий (ВСА), которые снабжают также переднюю и заднюю доли гипофиза. Нейросекреторные нейроны (H), тела которых лежат в гипоталамусе, секретируют гормоны в венозную систему. Ж - третий желудочек головного мозга. (По Wilson. 1979.) | 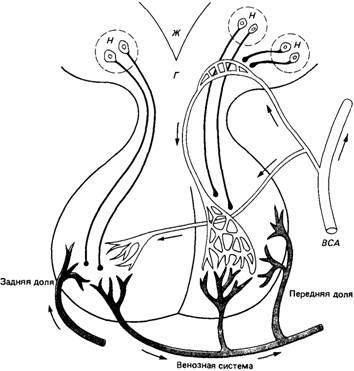
|
сахарный диабет, к симптомам которого относятся чрезмерное мочеотделение и вследствие этого жажда. Таким образом, антидиуретический гормон играет важную роль в сохранении воды.
К другим сохраняющим воду механизмам относятся повышение ее реабсорбции в тонком кишечнике - а следовательно, меньшая потеря с калом - и уменьшение количества поедаемой пищи. Из-за того что непереваренные остатки пищи и отходы метаболизма должны быть выведены, некоторая потеря воды неизбежна, но ее можно снизить, уменьшив количество потребляемой пищи. Лабораторные исследования показывают, что потери воды у голубей, лишенных корма, составляют лишь четверть нормального уровня (McFarland, Wright, 1969). Кроме того, по некоторым данным, голуби могут контролировать потери воды при дыхании (Wright, McFarland, 1969). Потерю воды на терморегуляцию иногда можно снизить определенным поведением - поисками прохладного места и снижением теплопродукции, вызываемой физической нагрузкой и потреблением пищи. Когда верблюду не хватает воды, он дает температуре тела повыситься и днем сохраняет тепло в жировых тканях горба. Во время холодной ночи в пустыне это тепло излучается без всякой потери воды. В противоположность распространенному мнению верблюды не хранят в горбах воду, хотя при метаболизме жировой ткани, разумеется, выделяется некоторое количество воды (Schmidt-Nielsen, 1964).
Животные обезвоживаются не только вследствие дегидратации клеток, но и за счет уменьшения объема внеклеточной жидкости. Геморрагия и другие формы кровопотерь не меняют осмотического баланса, но утраченная жидкость должна быть возмещена. Для обнаружения такой потери у животных имеются разные механизмы. В ответ на снижение объема крови, протекающей через почки, вырабатывается гормон ренин. Он поступает в кровь и там стимулирует синтез другого гормона - ангиотензина, который оказывает два основных действия: во-первых, поддерживает нормальное кровяное дав-
|
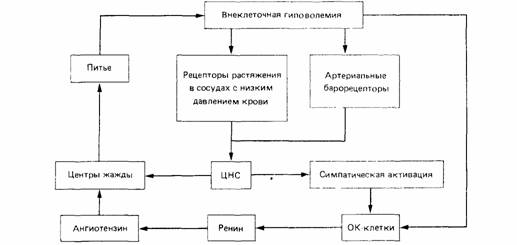
| Рис. 15.7. Внеклеточная жажда. Гиповолемия (потеря внеклеточного объема), обнаруживаемая рецепторами кровяного давления, вызывает секрецию ренина околоклубочковыми клетками (ОК). Ренин превращается в гипотензин, который стимулирует питье. (По Fitzsimons, 1971.) |
ление - а следовательно, регулирует кровообращение, - во-вторых, служит мощным возбудителем жажды (животные, получившие ничтожные количества ангиотензина. прерывают любую свою деятельность и ищут воду (Fitzsimons. 1976). Главные компоненты сложных механизмов внеклеточной регуляции жажды показаны на рис. 15.7.
Поддержание водного баланса тесно связано с терморегуляцией и питанием. Многие животные при обезвоживании сильно снижают потребление пищи, что очень способствует сохранению воды, так как принятие пищи, как мы видели, обычно вызывает значительную потерю воды путем экскреции. Основные механизмы сохранения воды показаны на рис. 15.8. Важно иметь в виду, что способ ее сохранения некоторым образом невыгоден для животного. Он может мешать нормальной терморегуляции или снижать поступление энергии. Различные механизмы по-разному важны для разных видов, что зависит от нормальных экологических условий жизни животного. Так. верблюд, чтобы сохранить воду, жертвует постоянной температурой тела, а голубь отказывается от еды. чтобы эта температура поддерживалась примерно на одном уровне (см. рис. 15.2).
На первый взгляд система регуляции водного баланса служит наглядным примером гомеостаза: животное обнаруживает отклонения от нормальных количеств (объема) и концентрации (осмолярности) внеклеточной воды и принимает меры для исправления положения, поглощая воду или снижая ее потери посредством тех или иных механизмов. Однако ситуация не гак проста из-за взаимодействия с другими системами. Например, часто животное пьет, когда оно не обезвожено.
Дата добавления: 2016-06-18; просмотров: 2889;
Поиск по сайту
Узнать еще
- II. Поведение веществ в магнитном поле.
- АГРЕССИВНОЕ ПОВЕДЕНИЕ
- Агрессивное поведение. Основные положения
- АДДИКТИВНОЕ ПОВЕДЕНИЕ НЕСОВЕРШЕННОЛЕТНИХ
- АФФИЛИАТИВНОЕ ПОВЕДЕНИЕ
- Бестактное поведение руководителя
- Блок 9. «Организационное поведение», «Управление персоналом»
- Влияние окружающей среды на поведение

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
