Координация и ориентация
В этой главе будет рассказано о пространственных отношениях животного с его внешней средой. В ответ на изменения среды животные должны координировать и ориентировать свои движения по отношению к внешним стимулам. Сначала мы рассмотрим координацию, а затем ориентацию и закончим обсуждением навигации — самой сложной формы пространственной ориентации.
Координация
Как показано в гл. И, эффективная координация движения и локомоции зависят от информации, получаемой ЦНС о положении, напряжении и других состояниях мышц; эта информация поступает от внутренних органов чувств - мышечных веретен, сухожильных органов и суставных рецепторов. Кроме передачи информации об относительном положении конечностей и других органов движения эти органы чувств позволяют животным давать рефлекторные ответы на изменения, вызываемые внешними агентами или самим движением животного.
Рефлекторное поведение - самая простая форма реакции на стимулы. В норме рефлексы бывают автоматическими, непроизвольными и стереотипными. Внезапное изменение напряжения мышцы может вызвать автоматическую смену позы, а резкое изменение уровня освещенности - рефлекторное отстранение. Рефлексы могут быть относительно локальными, т.е. затрагивать одну конечность или какую-либо другую часть тела, как показано на рис. 14.1, но иногда в рефлекторную реакцию вовлекается все животное. Например, реакция вздрагивания у человека требует рефлекторной координации многих мышц. Точно так же рефлекс отстранения у беспозвоночных - полихет и различных моллюсков - охватывает все тело. Как было показано в гл. 11, такие рефлексы запускаются гигантскими нервными волокнами, которые очень быстро передают сигналы всем участвующим в ответе мышцам, так что они сокращаются сразу и одновременно.
Рефлексы у позвоночных в норме автоматические, однако нередко они подвержены различным воздействиям через центральные синапсы (рис. 14.1). Так, с рефлексами позы часто взаимодействуют другие, несовместимые с ними рефлекторные механизмы. Если в двух рефлексах используются одни и те же мышцы, то эти рефлексы несовместимы, т.е. не могут протекать одновременно. Такие пары рефлексов несовместимы также неврологически в том смысле, что стимуляция одного рефлекса тормозит другой. Торможение обычно бывает реципрокным и составляет самую элементарную форму координации, когда одна активность полностью подавляет другую или подавляется ею. Этот тип реципрокного торможения характерен для ходьбы и других видов локомоции.
Вообще мышечная координация определяется двумя главными процессами - центральной и периферической регуляциями. При центральной регуляции головной мозг дает точные команды, которые выполняются соответствующими мышцами. К такому типу, по-видимому, относится координация глотательных движений у млекопитающих (Doty, 1968). Центральная регуляция важна для координации многих выученных движений, требующих
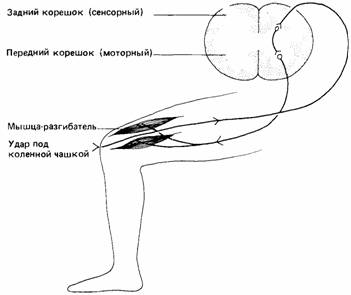
| Рис. 14.1. Схема простого коленного рефлекса. Удар под коленной чашкой стимулирует рецепторы в сухожилии, которые посылают сигналы в спинной мозг. Сигналы передаются двигательным нервам, управляющим мышцами-разгибателями, которые рефлекторно сокращаются, подбрасывая ногу. |
быстрой мышечной активности. Так, например, каракатица Sepia ловит мелких ракообразных двумя вытягивающимися щупальцами (рис. 14.2). Регуляция нападения делится на две фазы. Первая представляет собой направляемую зрением систему, в которой добыча приводится бинокулярно в фокус, и движения каракатицы следуют за движениями жертвы таким образом, что ошибка «прицеливания» сводится к нулю. Когда фаза слежения завершена, щупальцы внезапно выбрасываются и схватывают жертву. Завершающая фаза такая быстрая (около 30 мс), что времени на зрительный контроль движения не остается. Это можно показать, выключив свет во время выбрасывания щупалец: все равно жертва будет поймана. Но если в это время добыча переместится, то каракатица ее не поймает; значит, это головоногое не способно корректировать свой бросок, используя зрительную обратную связь (Messenger, 1968).
Локомоторные ритмы часто генерируются центральными механизмами. Отведение электрической активности моторных и вставочных нейронов у насекомых выявило центральные регулирующие ритмы при ходьбе (например, Pearson, Iles, 1970; Burrows, Horridge, 1974), плавании (например, Kennedy, 1976) и полете (например, Wilson, 1968). Однако на точную структуру поведения часто влияют рефлексы и обратная связь с периферии. Так, устранение проприоцептивных сигналов снижает частоту взмахов крыльев у летящей саранчи, а действие ветра усиливает разряд мотонейрона (Wilson, 1968). По-видимому, периферические стимулы действуют на быструю ходьбу таракана меньше, чем на медленную ходьбу его родичей палочников (Carausius) (Wendler, 1966).
Периферическое управление координацией происходит посредством органов чувств, располагающихся в мышцах и
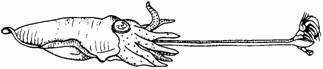
| Рис. 14.2. Каракатица, хватающая креветку. |
|
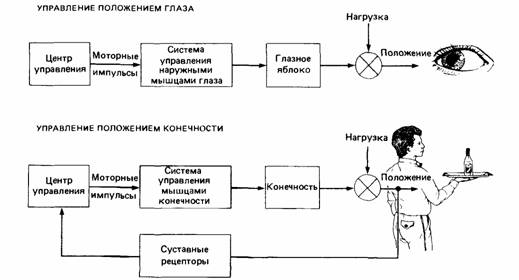
| Рис. 14.3. Центральная и периферическая регуляция движения. Движение глазного яблока (вверху) управляется центральной нервной системой. В норме глазное яблоко не подвергается нагрузке, поэтому подробные моторные команды, идущие от мозга, выполняются быстро и точно. Если же на глазное яблоко создать нагрузку (например, надавить пальцем), то его смещение не корректируется. Периферический контроль важен при движении конечностей (внизу), потому что они часто бывают нагружены. Изменение в положении конечности регистрируется сенсорными рецепторами, и по обратной связи о нем сообщается управляющему центру, который корректирует смещение. |
других частях тела; эти органы чувств посылают информацию в мозг и тем самым влияют на его команды мышцам (рис. 14.3). Периферическая регуляция обычно осуществляется совместно с центральной. Например, при координации плавательных движений у рыб головной мозг посылает ритмические сигналы, которые волнами проходят по спинному мозгу, координируя ритмические движения плавников и хвоста. У кошачьей акулы (Scyliorhinus) при перерезке всех нервов, ведущих от мышц к головному мозгу, этот ритм исчезает, но если сохранить некоторые нервы, он продолжается. Следовательно, некоторая периферическая обратная связь, по-видимому, необходима для запуска центрального ритма (Gray, 1950). У хрящевых рыб (Chondrichthyes), в том числе у кошачьей акулы и у всех других акул и скатов, плавники производят небольшие независимые ритмические движения, но у костистых рыб (Teleostei) они при некоторых обстоятельствах ударяют с разной частотой. Хольст (Hoist, 1939, 1973) показал, что ритмы разных плавников могут влиять друг на друга,-это свойство он назвал «относительной координацией». Иногда ритм одного плавника подстраивает под себя ритм другого и доминирует над ним так, что движения происходят в фазу. В других случаях амплитуды движений плавников суммируются, уменьшаясь, когда плавники движутся со сдвигом по фазе и увеличиваясь при совпадении фаз.
У рыб и амфибий локомоция, по-видимому, находится под контролем главным образом эндогенных спинальных ритмов. Нет данных о специальном моторном контроле со стороны переднего мозга. Его удаление у рыб не меняет их позы или локомоции (Bernstein, 1970). У лягушек и жаб это вызывает общее снижение спонтанных движений, но электрическая стимуляция их переднего мозга не приводит к специфическим моторным эффектам. Средний мозг играет некоторую роль в
моторной регуляции у рыб и амфибий. Как было указано в гл. 13. электрическая стимуляция тектума у жаб вызывает поворот головы, а также схватывание и проглатывание корма. У эволюционно более продвинутых позвоночных высший отдел головного мозга в большей степени способен контролировать движения, но автоматические компоненты локомоции по-прежнему регулируются прежде всего его стволом и спинным мозгом.
У млекопитающих кортикоспинальный тракт является самым важным проводящим путем, участвующим в произвольной регуляции движения. Он начинается в двигательной коре и проходит через средний мозг и мозговой ствол к спинному мозгу. Эта система, иногда называемая пирамидной, имеется у всех млекопитающих, за исключением очень примитивных однопроходных (например, утконоса и австралийской ехидны). У сумчатого кистехвостого кускуса (Trichosaunis) пирамидные аксоны доходят только до грудного отдела, где они иннервируют передние конечности. Задние конечности иннервируются экстрапирамидной системой.
Экстрапирамидная система включает все двигательные пути, не входящие в кортикоспинальную или пирамидную системы. Она считается более примитивной системой, чем пирамидная. У животных со слаборазвитой корой больших полушарий или вовсе лишенных ее базальные ганглии экстрапирамидной системы являются основными центрами моторного контроля. Они особенно хорошо развиты у птиц, у которых практически нет коры, но стриатум крупнее, чем у млекопитающих. Таким образом, в ходе эволюции кора больших полушарий, по-видимому, стала вторым источником двигательной координации, действующим через пирамидную систему.
У обезьян нейроны в разных частях кортикоспинальных путей меняют характер своей импульсации при произвольных • движениях глаз, передних и задних конечностей (Evarts. 1968). Искусственная стимуляция двигательной коры вызывает реакции отдельных мышц или отдельных моторных единиц в мышце. Более обширная стимуляция приводит к отдельным
|
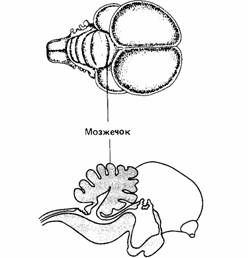
| Рис. 14.4. Головной мозг голубя с крупным мозжечком |
движениям целой конечности. Можно нанести карту на поверхность коры в соответствии с теми частями тела, которые движутся в ответ на ее электрическую стимуляцию. Можно построить такую же карту для сенсорной коры, на которой разные части тела представлены по-разному в соответствии с их сенсорным значением для данного вида. Произвольные выученные движения вызываются корковыми пирамидными нейронами, а рефлекторное поддержание двигательных реакций и позы управляются расположенными поблизости экстрапирамидными нейронами. Число пирамидных нейронов у человека оценивается в один миллион (Prosser, 1973).
Другой важной для координации частью головного мозга позвоночных является мозжечок (рис. 14.4). Он участвует в позных рефлексах или в управлении движением косвенно, являясь контролером и координатором нервной активности при ориентации, сохранении равновесия и других тонких реакциях, связанных с регуляцией движения. Основная нейронная организация мозжечка одинакова во всех классах позвоночных - он претерпел меньше эволюционных изменений, чем любая другая часть головного мозга. Мозжечок получает информацию от органов зрения,
слуха, осязания, равновесия, о состоянии мышц и суставов. Он связан также с двигательными областями коры большого мозга. Кроме связей через таламус с двигательной корой он имеет двусторонние связи с сенсорными областями коры, которые играют важную роль в поддержании позы. Вертикальная поза не может сохраняться без зрительной, вестибулярной и проприоцептивной информации. Мозжечок сочетает зрительную и вестибулярную информацию о равновесии с информацией о сокращении соответствующих мышечных веретен и посылает необходимые команды мышцам, в особенности по эфферентным гамма-волокнам (см. гл. 11). Таким образом, мозжечок осуществляет модулирующий тонкий контроль над мышечными сокращениями, связанными с поддержанием позы и со сложными координационными движениями, которые требуются для локомоции.
Дата добавления: 2016-06-18; просмотров: 3978;
Поиск по сайту
Узнать еще
- I. Ориентация формы в среде.
- Дискоординация родовой деятельности (гипертоническая дисфункция, некоординированные схватки)
- И «спортивная ориентация»
- Координация в олигополиях: способствующие и ограничивающие факторы
- КООРДИНАЦИЯ ДВИЖЕНИЙ
- Координация деятельности центральной нервной системы
- Координация и выполнение поиска опасных веществ и устройств

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
