Мотивационные системы
Требования гомеостаза, как мы видели, ставят определенные задачи перед поведением животных. В каждый данный момент животное должно оценивать свое внутреннее состояние, добавлять к этому свои знания о вероятных будущих нуждах и о тех новых нуждах, которые возникнут в ходе той или иной деятельности, и затем выбирать, что делать дальше.
Традиционное понимание мотивации основано на принципе простой обратной связи. Изменение во внутреннем состоянии животного воспринимается головным мозгом и побуждает к определенному поведению (создает драйв). Такое поведение бывает аппетитивным и консумматорным. Аппетитивное поведение включает в
себя поиск подходящих внешних стимулов; когда они найдены, наступает консумматорная активность, например потребление пищи или воды. Консумматорное поведение ведет к снижению драйва - или непосредственно, или путем уменьшения внутренних или внешних стимулов, которые его вызвали. Тогда консумматорное поведение также прекращается. Например, обезвоживание тканей тела ощущается головным мозгом и приводит к возникновению драйва жажды; он побуждает животное искать воду (аппетитивное поведение); когда вода найдена, животное пьет (консумматорное поведение). Питье может снизить жажду непосредственно, т. е. через кратковременные механизмы насыщения, такие, как ощущение воды во рту или тяжесть от воды в кишечнике (Rolls, Rolls, 1982). Кратковременные механизмы насыщения дополняются или обходятся у некоторых видов животных (Rolls, Rolls, 1982) поступлением воды в кровоток, что ослабляет дегидратацию, вызвавшую драйв жажды. Таким образом, животное перестает пить или под действием кратковременных механизмов насыщения, или благодаря тому, что состояние его водного баланса больше не вызывает жажды.
Термин драйв введен Вудвортом (Woodworth, 1918) как альтернатива выдвинутому Мак-Дугаллом (McDougall, 1908) понятию инстинкта (см. гл. 20). Вудворт различал в мотивации способность возбуждать энергию (драйв) и направлять деятельность. Первичные драйвы возникают из потребностей тканей, а вторичные - из приобретенных навыков. Сходные представления о драйве развивали первые этологи. Например, Лоренц (Lorenz, 1950) излагает этологическую концепцию в виде трех последовательных процессов: 1) накопление специфической для данного действия энергии, вызывающее аппетитивное действие; 2) аппетитивное поведение, направленное на достижение стимульной ситуации, которая активирует врожденный запускающий механизм, и 3) приведение в действие запускающего механизма и разряд эндогенной активности в консумматорном действии. Лоренц постулировал, что «некоторый вид энергии, специфичный для определенной активности, сохраняется, пока эта активность не наступила, и потребляется при ее осуществлении».
Основную идею драйва как побуждения к определенным действиям разделяли этологи и разные школы психологии животных в США. В течение 50 лет со времени его возникновения понятие драйва неоднократно обсуждалось и анализировалось на квазифилософском уровне. Споры шли вокруг вопросов: присуща ли драйвам целенаправленность (Thorpe, 1956; Peters, 1958), общий или специфический у них характер (см. Hinde, 1970; Bolles, 1967), можно ли считать, что драйв обеспечивает поведение энергией (Bolles, 1967; McFarland, 1971). В последние годы возникла тенденция отказаться от концепции драйва по причинам, изложенным ниже.
Классический взгляд на голод и жажду как на гомеостатические драйвы предполагает, что потребление пищи и воды — ответ на обнаруженные изменения в физиологическом состоянии животного. Считается, что эти действия управляются отрицательной обратной связью, потому что они служат уменьшению физиологических отклонений, вызвавших такое поведение. Однако прием пищи и воды не только ответ на физиологические изменения, но часто и предварение их. У многих животных имеется определенный «режим» питания, повторяемый при постоянных условиях ежедневно. Как люди испытывают чувство голода в определенные часы, так и у животных стремление к принятию пищи может определяться временем суток. Когда внешняя среда в разные дни мало меняется, у животных быстро устанавливается суточный порядок активности, и они едят в определенные часы даже при постоянном наличии пищи. Физиологические процессы могут подстраиваться к этому порядку. У человека, например, печень может прекратить мобилизацию гликогена как раз перед едой. Это ведет к падению уровня сахара в крови «в предчувствии» его повышения после переваривания пищи. Опыты показали, что такие физиологические приспособления могут вырабатываться условно-рефлекторно применительно ко времени дня.
Мы видели, как отдаленные последствия принятия пищи приводят к усилению жажды. Многие животные пьют не от такой жажды, а заранее, тем самым предотвращая обезвоживающее действие приема пищи (Fitzsimons, Le Magnen, 1969). Подобным же образом, как уже говорилось, терморегуляция часто связана с потерей воды, но некоторые животные пьют не в ответ на вызванную ею дегидратацию, а заранее, запасая в результате воду для регуляции температуры тела (McFarland, 1970a). Так, было обнаружено (Budgell, 1970a), что египетские горлицы (Streptopelia risoria), не получавшие воды в течение двух дней при разных температурах, выпивали одно и то же количество воды в экспериментальной камере при температуре 20°С. Однако горлицы, лишенные воды в течение двух дней при одной и той же температуре (20°С), затем выпивали разные ее количества при разных температурах (рис. 15.11). Аналогичные результаты, полученные на крысах (Budgell, 1970), показывают, что эти животные пьют, непосредственно реагируя на изменения температуры среды, еще до наступления какой-либо тепловой дегидратации. Термин предваряющая связь применим к ситуациям, когда последствия поведения, вызываемого обратной связью, предвосхищаются и принимаются соответствующие меры по предотвращению физиологических отклонений (McFarland, 1971; Toates, 1980).
Причины какой-либо активности и ее физиологические последствия не всегда однозначно соответствуют друг другу. В некоторых случаях, например при поведенческой терморегуляции, активность животного может приводить лишь к изменению температуры тела. Но чаще поведение животного сказывается на его состоянии разносторонне. Так, принятие пищи изменит множество физиологических процессов в зависимости от состава поедаемых продуктов. Последствия такого поведения называют амбивалентными.
Мы с Сибли (Sibly) показали в 1972 г., что для уравновешивания амбивалентных последствий поведения животные должны быть способны к адаптивному контролю, меняющему свойства их регуляторных
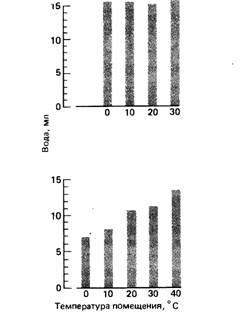
| Рис. 15.11. Действие температуры в помещении на питье у горлиц. Вверху: влияние температуры во время лишения воды на последующее питье. Внизу: влияние температуры во время питья. (Данные Budgell, 1970a.) |
механизмов в соответствии с условиями среды. Известно много примеров такого адаптивного контроля. Так, я обнаружил (McFarland, 1971), что горлицы научались изменять свою оценку ближайшего гидратирующего действия выпиваемой воды после испытанных ими переходов с чистой воды на соленую, и наоборот. Мы уже видели, как крысы научаются избегать определенную пищу или выбирать ее на основе физиологических последствий, наступающих через несколько часов после еды (см. также гл. 18). Было показано (Richter, 1943), что после хирургического удаления определенных физиологических регуляторов животные стараются поддерживать гомеостаз с помощью поведенческих реакций. Такого рода компенсаторному поведению научались крысы, помещенные в подходящую среду после удаления надпочечников, щитовидной, паращитовидной и поджелудочной желез. Иными словами, животные, лишенные физиологической регуляции, обращались к вспомогательным средствам.
|
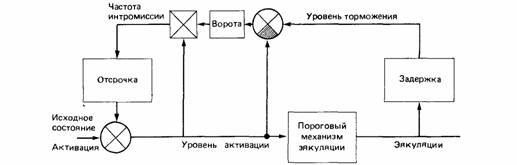
| Рис. 15.12.Эта модель регуляции брачного поведения у самцов крыс в принципе мало отличается от моделей, предложенных для регуляции еды или питья. (По McFarland, Nunez, 1978.) |
Чтобы понять физиологические и поведенческие механизмы гомеостаза, недостаточно представлений о простой отрицательной обратной связи. Поддержание физиологического состояния в узких пределах достигается сочетанием отрицательной обратной связи, предваряющей связи и адаптивного контроля.
Традиционно различались гомеостатические стороны поведения - питание, питье и терморегуляция - и негомеостатические виды активности, например агрессия и половое поведение. В последние годы правомерность такого деления оспаривается (см., например, Hogan, 1980; Davis, 1980). Регуляция полового поведения в своей основе не отличается от регуляции питания или питья (McFarland, Nunez, 1978; Toates, 1980). Принципы контроля в целом одинаковы и могут быть представлены моделями, использующими одни и те же термины и представления (рис. 15.12).
Физиологические процессы, определяющие поведение, находятся в сложном взаимодействии. В возникновении, поддержании и прекращении простой на первый взгляд активности (например, питья) может участвовать несколько разных факторов (Rolls, Rolls, 1982). При питании ситуация гораздо сложнее (Booth, 1978). Чтобы разобраться в этой сложности, ученые, изучающие поведение, обратились к применяемым инженерами для описания и анализа сложных машин методам теории систем управления. Создав количественные модели различных компонентов системы (например, управляющей питьем), можно получить ее компьютерную имитационную модель как целого. Такая модель может служить для количественных предсказаний, проверяемых опытами, причем результаты этих опытов используются для уточнения гипотез и создания на этой основе все более совершенных моделей. Теория систем управления применима к разным типам поведения, в том числе к питанию, питью, терморегуляции и половому поведению (McFarland, 1971, 1974; Booth, 1978; Toates, 1975, 1980).
По сравнению с точностью и строгостью теории систем управления концепция драйва расплывчата и запутанна. Кроме того, новый подход вскрыл концептуальные проблемы, которые наглядно демонстрируют нежизнеспособность понятия драйва. Две такие проблемы были точно указаны Хайндом (Hinde, 1959, 1960), но необходимые для рассмотрения их следствий теоретические представления тогда еще не были приняты в области поведения животных. Одна из ошибок (Hinde, 1959) состоит в том, что драйвы можно рассматривать как однозначные переменные величины. Думая о драйве голода, мы представляем себе величину, измеримую по одной шкале. Так, мы думаем, что животное слегка или же сильно голодно. Но, как уже говорилось, голод имеет много аспектов. Животное может испытывать специфический голод в отношении, скажем, соли или тиамина. У него может быть недостаток белка или
легкодоступной энергии. Эти особенности могут влиять на пищевое поведение, поэтому неправильно говорить о голоде как об одномерной переменной. Альтернативная формулировка (McFarland. Sibly, 1972) представляет голод и другие так называемые драйвы как векторы. Такой подход послужил основой для изображения мотивационных систем в виде пространственных моделей, к чему мы вернемся ниже.
Вторая ошибка (Hinde. 1960) - представление о драйве как об источнике энергии для поведения. Хотя эта точка зрения была очень влиятельна в психологии и этологии, она создает трудности, как только вопрос выходит за рамки общей аналогии. Многие прежние психологи механистического направления основывали свои взгляды на неприменимых в данном случае физических понятиях силы, мощности и энергии. Как и первые этологи, они приравнивали драйвы к энергии (см. выше), считая, что некая мотивационная энергия накапливается при пищевой, питьевой, половой депривации и что эта энергия, или драйв, определяет интенсивность последующею поведения. Основная трудность здесь состоит в том. что энергия в физическом смысле представляет собой способность и сама по себе не может быть причиной действия (см. McFarland, 1971). Исходя из неверных аналогий, теории драйва стали неприменимыми и противоречивыми (Bolles, 1967).
Третья важная проблема, связанная с понятием драйва, возникла при попытках классификации. Некоторые психологи стремились выделить драйв для каждого вида поведения; этот подход присущ ранним этологическим теориям. Так. можно было бы постулировать сексуальный драйв, определяющий половую энергию, но можно и разделить его на драйв ухаживания, драйв спаривания и драйв эякуляции. Возникает вопрос: сколько должно быть драйвов? Часть психологов высказывалась в пользу единого общего драйва, но и это предложение оказалось неудовлетворительным (Hinde, 1970; Bolles, 1967). Если же отказаться от понятия драйва и сосредоточить внимание на изменениях мотивационного состояния, лежащих в основе поведения, то можно перейти к выявлению связанных с этим факторов (McFarland, 1974).
Дата добавления: 2016-06-18; просмотров: 2110;
Поиск по сайту
Узнать еще
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2.1 ПЕРВЫЙ ЗАКОН НЬЮТОНА. ИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА.
- I.2.1 ПОЛНАЯ И ВНУТРЕННЯЯ ЭНЕРГИЯ СИСТЕМЫ. ТЕПЛОТА И РАБОТА
- I.3. Антитела системы АВ0
- I.5.4 НЕИНЕРЦИАЛЬНЫЕ СИСТЕМЫ ОТСЧЁТА. СИЛЫ ИНЕРЦИИ
- II. ЭЛЕКТРИЧЕСКИЙ ДИПОЛЬ. ДИПОЛЬНЫЙ МОМЕНТ СИСТЕМЫ ЭЛЕКТРИЧЕСКИХ ЗАРЯДОВ
- III. Избирательные системы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
