Принцип реафферентации
Высокоразвитая система ориентации должна быть способна отличать стимулы из внешнего мира от стимуляции, создаваемой самим животным. Так, например, что касается зрения, то у человека перемещение объектов во внешнем мире вызывает движение изображения по сетчатке, которое мы воспринимаем. Однако произвольное движение глаз тоже вызывает перемещение изображения на сетчатке, но оно уже не воспринимается. Каким-то способом мозг отличает перемещение изображения по сетчатке, независимое от животного, от перемещения, вызываемого движением самого глазного яблока.
Для объяснения этого явления предложены две теории - оттока и притока. Согласно теории оттока, основы которой заложил Гельмгольц (Helmholtz, 1867). команды глазным мышцам о движении глазного яблока сопровождаются параллельными сигналами, идущими к компаратору в головном мозгу. Здесь они сравниваются с приходящими зрительными сигналами (рис. 14.10). Согласно теории притока, созданной Шеррингтоном (Sherrington, 1918), рецепторы наружных глазных мышц посылают сообщения в мозговой компаратор при любых движениях глаз (рис. 14.10). В обеих теориях компаратор оценивает оба приходящих
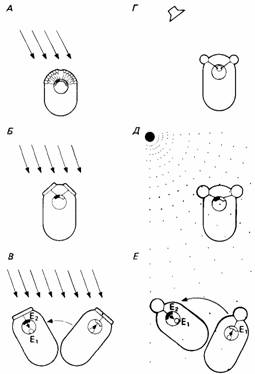
| Рис. 14.8. Схематическое изображение некоторых основных принципов сенсорной ориентации. А. Направление стимуляции (например, снега) регистрируется растром сенсорных рецепторов. Б. Направление регистрируется посредством сравнения одновременной стимуляции двух рецепторов. В. Имеется только один рецептор, и животное проводит последовательные сравнения, передвигая туловище. Г. Время прихода стимула (например, "звуковых волн) сравнивается двумя рецепторами. Д. Градиент стимуляции (например, химической) peгистрируется посредством сравнения данных, получаемых двумя рецепторами. Е. Градиент регистрируется одним рецептором, когда животное движется, обследуя разные точки пространства. (Из The Oxford Companion to Animal Behaviour, 1981.) |
сигнала и определяет, соответствуют ли зрительные сигналы тому движению, которое следовало ожидать на основании второго сигнала. Если сигналы не соответствуют друг другу, значит, какая-то часть движения должна была быть вызвана внешними причинами.
Наружные глазные мышцы содержат
|
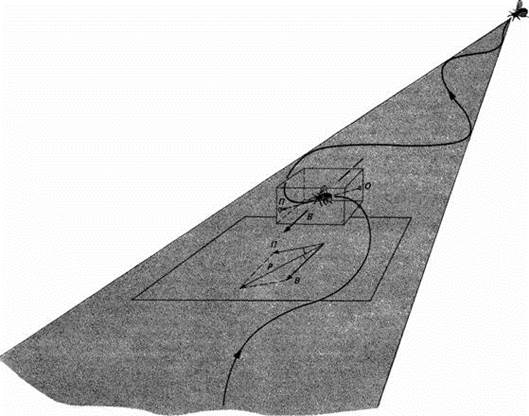
| Рис. 14.9.Самец бабочки, летящей против ветра на феромон, выделяемый самкой. На направление полета П влияет направление ветра В, и возникает результирующий маршрут Р. Угол передвижения - это угол между направлением ветра В и направлением Р по отношению к земле. |
мышечные веретена, и их существование, казалось бы, подтверждает теорию Шеррингтона. Однако, по-видимому, эти мышцы не участвуют в подаче сигналов о положении глаза (Merton, 1964; Howard, Templeton. 1966). Для доказательства того, что в глазных мышцах нет рецепторов положения, Гельмгольц (Helmholtz, 1867) пользовался данными по механическим манипуляциям с глазным яблоком и кажущимся движениям, вызываемым попытками двигать глазом, когда наружные глазные мышцы парализованы.
Как хорошо известно, при смещении глазного яблока в глазнице нажимом на него пальцев зрительная ось сдвигается (что можно заметить по раздвоенному изображению); она остается сдвинутой, пока длится нажим. Глазное яблоко не давит на палец чтобы занять свое прежнее положение, как можно было бы ожидать, если бы в регуляции положения глаза участвовали мышечные веретена (McFarland, 1971). Кроме того, при смещении глазного яблока пальцем движение ощущается, чего не должно бы быть по теории притока. Мышечные веретена возбуждались бы при любом движении глазного яблока, и тогда компаратор в мозгу компенсировал бы движение изображения по сетчатке. Поэтому факты, по-видимому, говорят в пользу теории оттока.
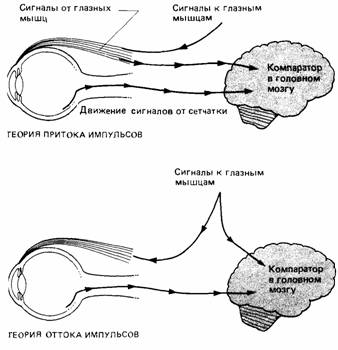
| Рис. 14.10. Теории притока и оттока сигналов (импульсов) при управлении движением глазного яблока. |
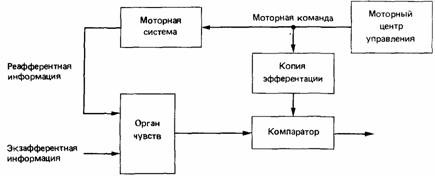
Теория оттока была развита и обобцена Хольстом и Миттельштедтом Holst, Mittelstaedt. 1950; см. также Holst, 1954). Согласно их принципу реафферентации, мозг отличает экзафферентную стимуляцию (вызываемую только факторами вне животного) от реафферентной, происходящей в результате движений тела животного). Моторные команды не только организуют мышечные движения, но и создают их нервную копию (копию "эфферентации»), соответствующую сенсорным сигналам, которые можно было ожидать при данном поведении животного. В результате мозг сравнивает копию эфферентации с приходящей сенсорной информацией (рис. 14.11). Вся реафферентная информация должна быть погашена копией эфферентации так, чтобы выход из компаратора был равен нулю. В то же время экзафферентная информация нe аннулируется, а передается компаратором в другую часть мозга.
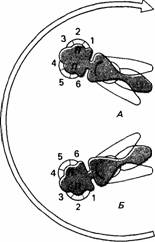
Хольст и Миттельштедт (Holst, Mittelstaedt, 1950), показали, что если двукрылую ильницу (Eristalis) поместить в цилиндр с вертикальными полосами, у нее наблюдается типичный оптомоторный рефлекс, т. е. она поворачивается в направлении полос при вращении цилиндра. Такие рефлексы не возникают, если ильница движется сама по себе, хотя зрительная стимуляция при этом такая же. Если в опыте повернуть этому насекомому голову на 180 (рис. 14.12), то, как и следовало ожидать, оптомоторный рефлекс становится обратным. Но если ильница пытается двигаться сама, го начинает вертеться на месте, и ее движения, по-видимому, самовозбуждаются. Эти данные можно объяснить, исходя из теории реафферентации. В норме выходные сигналы компаратора определяют движение тела, а когда двукрылое движется само по себе, выход равен нулю и движения нет. Оптомоторный аппарат выдает экзафферентную стимуляцию, которая не аннулируется копией эфферентации, и ильница отвечает рефлексом. Когда голова ее повернута на 180 , экзафферентная стимуляция вызывает то же действие, что и раньше, но в обратном направлении. Однако в случае реафферентной стимуляции воспринимаемое движение имеет обратный знак и не вычитается из копии эфферентации, а прибавляется к ней, и в
|
| Рис. 14.11. Схема основной системы реафферентации. |
результате выход компаратора, который в норме должен быть сведен к нулю, возрастает. Чем больше животное реагирует, тем сильнее реафферентная стимуляция и тем больше усиливается реакция животного. В результате насекомое крутится на месте все быстрее и быстрее. Принцип реафферентации важен не только в отношении зрения, но также для контроля за положением конечностей, тела и т.д. Так, например, мы можем отличить движения руки, трясущей ветку дерева, от, возможно, таких же движений, возникающих, когда рука пассивно держится за ветку, раскачиваемую ветром.
Навигация
Самая сложная форма пространственной ориентации - это навигация. Она требует не только компаса, или дирекционального чувства, но и своего рода карты. Для иллюстрации рассмотрим опыт, проведенный Бейкером (Baker, 1981) для проверки того, может ли человек пользоваться магнитной информацией. Группа студентов была перевезена в небольшом автобусе от определенной начальной точки (дома) к некоторому секретному месту назначения. По приезде студенты должны были показать, в какой стороне находится их дом. Для точного ответа необходим внутренний магнитный компас, и Бейкер искал данные в пользу его наличия. Однако выполнение такой задачи требует также знания относительного положения обоих пунктов. Даже если бы студенты обладали точным магнитным компасом, они не могли бы указать, в каком направлении находится дом, без какой-нибудь карты.
При навигации важны три типа ориентации: 1) пилотирование, или прокладывание пути по знакомым ориентирам; 2) компасная ориентация, т. е. способность двигаться по определенному азимуту, не пользуясь ориентирами, и 3) истинная навигация, или способность направляться к цели - дому или месту выведения потомства, — не пользуясь ни ориентирами, ни компасом. В своих дальних миграциях птицы, вероятно, используют все три типа ориентации. Пердек (Perdeck,
|
| Рис. 14.12. Ильница Eristalis во вращающемся цилиндре. А. Нормальное положение головы. Б. Голова повернута на 180°. Л - левый глаз; П - правый глаз. (По van Hoist, 1954.) |
|
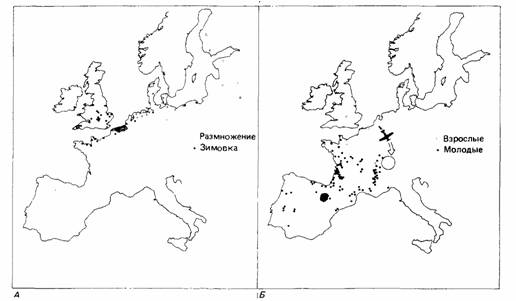
| Рис. 14.13. А. Отлов размножавшихся и зимовавших скворцов, окольцованных в Нидерландах. Б. Отлов взрослых и молодых скворцов, перевезенных из Нидерландов в Швейцарию и выпущенных там во время осенней миграции. (По Perdeck, 1958.) |
1958, 1967) отлавливал молодых и взрослых скворцов, когда они пролетали через Нидерланды при своей первой осенней миграции, кольцевал их и перевозил на самолете в Швейцарию, т. е. на 750 км юго-восточнее их обычного миграционного пути. В норме скворцы мигрируют от своих гнездовий на Балтике к зимовьям в Бельгии, южной Англии и северной Франции (рис. 14.13). После перемещения в Швейцарию молодых скворцов ловили в Испании и южной Франции (рис. 14.13); это говорит о том, что они сохраняли нормальное юго-западное направление миграции. Но взрослых скворцов снова ловили в местах их обычных зимовок. Таким образом, взрослые скворцы внесли поправку на свое перемещение, тогда как молодые сохранили компасную ориентацию, характерную для их местной популяции. Молодые животные многих мигрирующих видов в первый раз достигают мест своих зимовок на основе врожденной информации о направлении на цель и расстоянии до нее. Они не способны внести поправку на перемещение, потому что у них нет карты, необходимой для истинной навигации (Schmidt-Koenig, 1979).
Ориентация по окружающим объектам может играть важную роль в навигации, особенно при приближении к месту назначения. Классический пример этого приведен в работе А. Хаслера (A. Hasler, 1960) по миграции лосося. Тихоокеанские лососи (Oncorhynchus) вылупляются из икры в ручьях западной части США и Канады. Проходя стадию смолта, они скатываются вниз по реке в Тихий океан. Проведя два-три года в море, лососи становятся половозрелыми и возвращаются в тот же самый ручей, где родились. Путешествие к берегу, вероятно, происходит по солнечному компасу. Но дойдя до берега, они должны выбрать нужную реку и единственный из впадающих в нее ручьев. После многих лет работы Хаслер с сотрудниками установили, что на стадии смолта в рыбах «запечатлеваются» запаховые признаки их родного ручья (импринтинг). Во время обратного путешествия они отличают его воду от воды других речных притоков. В сущности они опознают ориентиры, но, поскольку запах
|
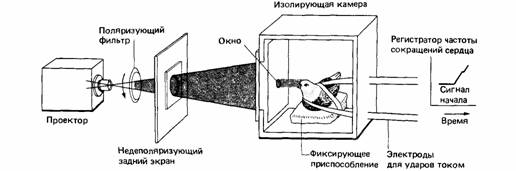
| Рис. 14.14. Прибор для выработки условного сердечного рефлекса у голубей при испытаниях их чувствительности к разным стимулам (в данном случае - к поляризованному свету). (По SchmidtKoenig, 1979.) |
места их рождения распространяется водой по всему пути миграции, молодому лососю не нужно запоминать его по дороге к океану. И действительно, если после периода импринтинга лососей перевести в другой ручей, они вернутся в свою родную речку, а не в ту, по которой мигрировали к океану.
Известно, что животные пользуются разного рода компасами, основанными на свойствах геофизической среды, например магнитным полем земли. Доказать, что определенное физическое свойство используется в качестве компаса, можно, продемонстрировав способность животного обнаруживать это явление и использовать его для ориентации в естественных условиях. Ввиду неуправляемой изменчивости природы проверять сенсорные способности лучше всего в лаборатории. Излюбленным методом исследователей навигации служит метод условных сердечных рефлексов (рис. 14.14). Этим и другими методами было показано, что голуби чувствительны к следующим стимулам:
Внешнее давление. Голубь чувствителен к изменениям атмосферного давления в пределах от 1 до 10 мм водяного столба, что соответствует изменению высоты менее 10 м (Kreithen, Keeton, 1974; Delius, Emmerton, 1978); благодаря такой сенсорной способности голубь обладает точным физиологическим альтиметром.
Инфразвук. Звук частотой менее 10 Гц называется инфразвуком. Человек не может слышать его, голуби же воспринимают частоты до 0,06 Гц. Инфразвук распространяется на очень большое расстояние, и такие его источники, как прибой, голуби могли бы использовать для навигации (Yodlowski et al., 1977; Kreithen, 1978).
Запах. Долго считалось, что у птиц слабое обоняние, но опыты с условными сердечными рефлексами (например, Henton et al., 1966; Shumake et al., 1966) подтверждают результаты применения физиологических методов и показывают, что обоняние у голубей достаточно тонкое, чтобы служить при навигации. Такие же данные получены для других видов птиц (Schmidt-Koenig, 1979).
Магнитный компас. Ученые долго считали, что энергия геомагнитных явлений слишком мала для того, чтобы животные могли ее воспринимать. Теперь известно, что это не так, и у многих видов показаны реакции на магнитные поля.
Хотя и раньше были указания на магнитную чувствительность у птиц (например, Merkel, Wiltschko, 1965), она долго вызывала сомнения, поскольку ее не удавалось показать в опытах с условными рефлексами на сердце. Но затем были получены положительные результаты не на привязанных, а на свободно передвигающихся голубях (Bookman, 1978).
Миниатюрные магниты обнаружены у бактерий, пчел и голубей. Хотя известно, что пчелы и голуби чувствительны к магнитным полям, не ясно, как магнитная информация воспринимается нервной системой. Имеются данные о том, что маг-
|
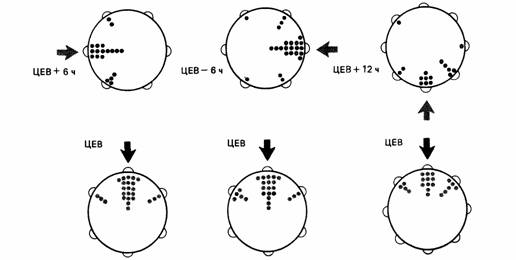
| Рис. 14.15. Результаты опытов на почтовых голубях со смешением времени суток. Птиц в круглой клетке обучали искать пишу в определенном азимутальном направлении. Учитывалось число клеваний кормушек, помещенных по окружности (каждая точка - один удар). ЦЕВ- центральноевропейское время. Стрелки - ожидаемое в условиях опыта направление. Верхний ряд - опыты. нижний одновременные контроли. (По Schmidt-Koenig, 1979.) |
нитные явления влияют на биологические часы пчел (Gould. 1980). У птиц чувство направления, по-видимому, связано со склонением магнитного поля Земли, и имеется предположение, что оно же важно для чувства карты у почтовых голубей (см. ниже).
Солнечный компас. Способность птиц использовать солнце как компас открыта Крамером (Kramer, 1951). Он приучал скворцов, находящихся в круглой клетке, искать корм в определенном азимутальном направлении. Все зрительные ориентиры были исключены, и видны были только солнце и небо. Птицы были способны находить нужный азимут в течение всего дня, что доказывает их способность делать поправку на движение солнца.
Биологические часы скворца (см. гл. 16.3) можно искусственно «перевести» (Hoffman. 1954), поместив птицу в светонепроницаемое помещение с искусственным фотопериодом. Шмидт-Кёниг (Schmidt-Koenig. 1958, 1960. 1961) в опытах с почтовыми голубями испытывал действие такого перевода на 6 ч вперед и на 6 или 12 ч назад. Результаты, представленные на рис. 14.15. говорят о том, что ориентация птицы в экспериментальном аппарате была точно такой же, какую можно было ожидать, пользуясь в качестве компаса солнцем.
Показано, что солнечный компас голубей достаточно точен для навигационных целей при условии, что в течение путешествия птицы время от времени делают поправочные измерения. Однако связь азимута солнца с местным временем пригодна прежде всего для опреде-, ления долготы, а с изменениями широты местонахождения наблюдателя меняется высота солнца над горизонтом. По некоторым данным опытов с инструментальными условными рефлексами, голуби могут достаточно точно определять изменения этой высоты. Имеются также данные, что они оценивают высоту солнца, измеряя скорее тени, а не само положение светила (McDonald, 1972, 1973). Действительно, размеры теней могут изменяться в шесть раз сильнее, чем высота солнца над горизонтом, но неизвестно, способны ли голуби использовать эту информацию вне помещения. Уайтен (Whiten, 1972) обучал голубей связывать высоту солнца с северным или южным на-
правлением от дома. Его опыты показывают, что голуби способны сопоставлять эту высоту с прохождением меридиана, однако нет данных об использовании такой способности при навигации.
Некоторые исследователи (например, Matthews, 1955, 1968; Pennycuick, I960) пробовали объяснить навигацию голубей, исходя из движения солнца. Теоретически информации достаточно для определения долготы (азимут солнца с поправкой на время) и широты (высота солнца над горизонтом в разное время), т. е. для построения эквивалента карты. Но это требует очень большой остроты зрения и точности увязывания движения солнца с временем. Большинство ученых сомневается в том, что голуби способны на такую точность (Schmidt-Koenig, 1979). Кроме того, способность летящего голубя определять высоту солнца никогда не была доказана, а между тем это весьма важно с точки зрения теорий навигации по солнцу.
Звездный компас. Если певчие птицы посажены в клетку в то время, когда обычно происходит их миграция, то они проявляют типичное дирекциональное миграционное беспокойство; ночью эта дирекциональность связана со звездами (Sauer, Sauer, 1955). Птицы были ориентированы под естественным звездным небом и под небом планетария. Методики регистрации миграционного беспокойства позднее были усовершенствованы (Emlen, Emlen 1966) с помощью аппарата, показанного на рис. 14.16. Эмлен (Emlen, 1967) обнаружил, что дирекциональность у индиговых овсянок (Passerina суапеа) одна и та же под естественным небом и под стационарным небом планетария. В то же время птицы следовали за смещением планетарного неба, и Эмлен (1972) обнаружил, что их ориентация была связана скорее с его вращением, чем с определенными созвездиями.
Наземному наблюдателю кажется, что звезды расположены на внутренней поверхности сферы, называемой небесной сферой. В каждый данный момент звезды расположены на небесной сфере определенным образом, причем образуемый ими рисунок движется так, что создает впечатление, будто эта сфера вращается. В действительности вращается Земля вокруг своей оси, проходящей через полюсы. Точка в северном небе, вокруг которой небесная сфера будто бы вращается, называется северным полюсом мира. Полярная звезда лежит очень близко к этому полюсу, и для практических целей ею можно пользоваться при определении направления на север от наблюдателя.
Эмлен (1972) показал, что овсянки, выращенные в неволе и совсем не видевшие неба, не могли ориентироваться во время миграции. Оперившиеся птенцы, помещенные до наступления осенней миграции под небо планетария, вращающееся вокруг Полярной звезды, показывали нормальную ориентацию на юг, А на север ориентировались птицы, находившиеся под экспериментальным небом планетария, которое вращалось вокруг звезды Бетельгейзе в созвездии Ориона, видимой наблюдателю в северных широтах в южной части неба. По-видимому, только что оперившиеся птенцы научались узнавать, какая часть неба вращалась меньше всего, и во время миграционного беспокойства ориентировались в противоположном направлении.
Методики условных сердечных рефлексов тоже были использованы для исследования восприятия птицами ночного неба. Получены данные, что кряквы (Anas platyrhynchos) научаются распознавать определенные расположения звезд (Wallraff, 1969). Хотя такие картины потенциально сходны по своей информации с картой, имеющиеся пока сведения говорят о том, что звезды используются только как компас (Schmidt-Koenig, 1979).
Поляризованный свет. В нормальном неполяризованном свете световые волны колеблются одинаково во всех плоскостях. В поляризованном свете более сильные колебания распространяются в одной плоскости. При рассеянии неполяризованного солнечного света молекулами атмосферы происходит поляризация, максимальная для света, рассеянного под углом в 90° к солнечным лучам, а это значит, что поляризация распределена в небе определенным образом, причем ее распределение меняется в соответствии с положением солнца.
|
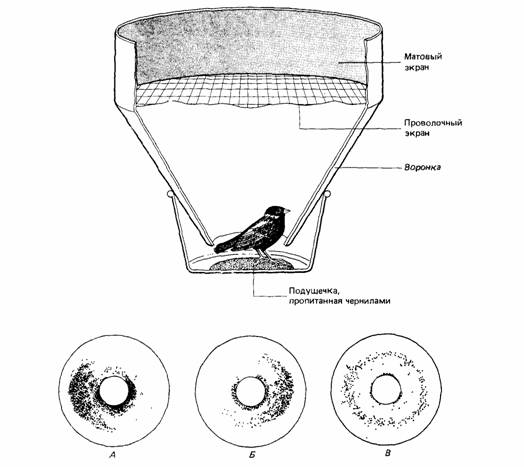
| Рис. 14.16. Экспериментальная клетка для измерения миграционного беспокойства. Пытаясь покинуть клетку, птица оставляет чернильные следы на промокательной бумаге, выстилающей воронку. Внизу - несколько примеров регистрации. (По Schmidt-Koenig, 1979.) |
Было обнаружено (von Frisch, 1967), что ориентация пчел, выражаемая их танцем (см. гл. 23), зависит от положения солнца, даже когда оно скрыто тучами. Оказывается, пчелам достаточно видеть только небольшую часть неба, причем нужная информация соответствует ультрафиолетовой части спектра. При пропускании ультрафиолетового света солнца через поляризующий фильтр ориентация танца пчелы менялась в соответствии с углом поляризации.
Опыты с условными сердечными рефлексами (см. рис. 14.14) ясно показывают, что голуби способны воспринимать вращение плоскости поляризации света (Kreithen, Keeton, 1974; Delius et al., 1976), но неизвестно, как они интерпретируют эту информацию. Хотя характер поляризации солнечного света может дать сведения о местоположении солнца даже тогда, когда небо покрыто тучами, по всей вероятности, голуби не пользуются своим солнечным компасом при таких условиях (Schmidt-Koenig, 1979).
ДЛЯ ЗАПОМИНАНИЯ
1. Координация движения подчиняется как центральному, так и периферическому управлению. Периферическое управление преимущественно задано заранее, тогда как центральное может включать в себя обратную связь (замкнутая петля) или же быть предсказующим (незамкнутая петля).
2. Пространственная ориентация простых животных основана на кинезах и таксисах, типы которых зависят от сенсорных способностей животного. Навигация как более сложная форма ориентации требует эквивалентов как карты, так и компаса.
3. Известно, что в навигации участвует множество разных сенсорных модальностей, но не совсем понятно, как координируется информация.
Рекомендуемая литература
Fraenkel G.S., Gunn D.L. (1940) The Orientation of Animals, Clarendon, Oxford (Dover Books, 1961).
Schmidt-Koenig K. (1979) Avian Orientation and Navigation, Academic Press, London.
Дата добавления: 2016-06-18; просмотров: 2265;
Поиск по сайту
Узнать еще
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. Этические принципы психолога
- II. Общие методические принципы в канистерапии
- II. Получение вращающегося магнитного поля и принцип действия АД.
- II. Принцип действия и режимы работы синхронной машины
- II. Электрическая схема и принцип действия.
- III. Принцип действия
- III. Принцип удовольствия

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
