Пространственная ориентация
Ориентация всего тела животного в пространстве может быть основана на очень простых принципах, но иногда включает и весьма сложные механизмы. Эти простые принципы легче всего наблюдать на некоторых видах беспозвоночных. Френкель и Ганн (Fraenkel, Gunn, 1940) предложили классификацию, основанную на работе более ранних авторов и послужившую отправным пунктом для последующих обсуждений и обзоров (например, Adler, 1971; Kennedy, 1945; Hinde, 1970).
Самая простая форма пространственной ориентации - это кинез, при котором реакция животного пропорциональна интенсивности стимула, но не зависит от его пространственных свойств. Например, мокрицы (Porcellio scaber) стремятся скапливаться во влажных местах под камнями или упавшим деревом. Они активно движутся при низкой влажности и менее активно - при высокой. В результате мокрицы проводят больше времени во влажной среде, и их высокая активность в сухих местах повышает их шансы попасть во влажные условия. Сходное поведение наблюдается у пескоройки - личинки речных миног, активность плавания у которой изменяется в зависимости от интенсивности освещения (Jones, 1955).
Тип кинеза, при котором скорость локомоции связана с интенсивностью стимуляции, называется ортокинезом. Другой тип, наблюдаемый у плоского червя Dendrocoelum lacteum, называется клинокинезом. При нем по мере усиления освещенности меняется скорость изменения направления (Ullyott, 1936; Fraenkel, Gunn, 1940; Hinde, 1970).
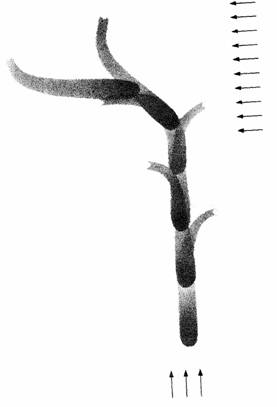
При многих типах ориентации, обычно объединяемых в группу таксисов, животное направляется прямо к источнику стимуляции или прямо от него. Например, когда личинка комнатной мухи (Musca domestica) прекращает питаться, она ищет темное место для окукливания. При этом она уползает прочь от источника света, что называют «отрицательным фототаксисом». У личинки на голове имеются примитивные глаза, регистрирующие изменения освещенности, но не способные давать информацию о направлении, в котором находится источник света. Когда личинка ползет, она поворачивает голову из стороны в сторону (рис. 14.5). Если свет слева ярче, чем справа, менее вероятно, что личинка повернет голову влево. Таким образом, она скорее поползет вправо, т. е. подальше от источника света. В ответ на повышение освещенности личинка усиливает скорость поворотов головы. Если свет над головой выключать каждый раз, когда она поворачивает голову вправо, и включать каждый раз при повороте ее влево, тогда личинка будет отворачиваться от освещаемой стороны, двигаясь по кругу вправо. Таким образом, хотя у животного нет дирекциональных рецепторов, оно способно к дирекциональной реакции. Такое же поведение наблюдается у одноклеточного эвглены с одним фоточувствительным «глазком» (Fraenkel, Gunn, 1940).
Ориентация путем последовательного сравнения интенсивности стимула требует поворотов. Обычно она называется клинотаксисом. У многих животных он проявляется в ответ на градиенты химических стимулов. Одновременное сравнение интенсивности стимулов, получаемых двумя
| 
|
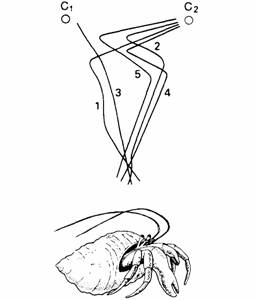
| Рис. 14.5. Клинотаксис у личинки мухи. Обратите внимание на движение головы из стороны в сторону. При перемещении находящегося сзади источника света в сторону личинка отворачивается от него. (По Mast, 1911.) | Рис. 14.6. Телотаксис: траектории движения крабов-отшельников при предъявлении им двух источников света (С1 и С2). Каждая часть пути направлена только к одному источнику. (По Fraenkel, Gunn, 1940.) |
или более рецепторами, дает возможность животному выровнять ее. В результате оно переходит к тропотаксису, который позволяет двигаться прямо к источнику стимуляции или от него. Например, мокрица Armadillidium vulgare, живущая под камнями или упавшими деревьями. проявляет положительный фототаксис после периодов высыхания или голода. Благодаря двум сложным глазам на голове животное способно двигаться прямо к источнику света. Если же один глаз зачернить, оно движется по кругу. Это говорит о том, что в норме оба глаза уравновешивают стимуляцию. При предъявлении двух световых источников мокрица часто начинает с того, что движется в среднем между ними направлении, а затем поворачивает к одному из них. Это происходит потому, что равная стимуляция обоих глаз достигается при движении либо точно между двумя источниками, либо прямо к одному из них. Отклонения от одного источника являются саморегулирующимися, а боковой свет игнорируется, потому что сзади и с боков глаза экранированы. В то же время отклонения от среднего маршрута между двумя широко расставленными источниками могут привести к потере контакта с одним из них.
Животные с глазами, дающими благодаря своему строению информацию о направлении света, способны к телотаксису форме дирекциональной ориентации, которая не зависит от одновременного сравнения стимуляции двух рецепторов. При двух источниках стимуляции животное движется к одному из них и никогда не избирает среднего направления: это показывает, что влияние одного из стимулов тормозится. Такой пример приведен на рис. 14.6.
Менотаксис - этотакая форма телотаксиса (Hinde. 1970), при которой ориен-
| Рис. 14.7. Солнечный компас: муравей (Lasius niger) возвращается в гнездо Г, когда солнце светит справа под углом около 90 к направлению движения животного. В точке X муравей был задержан на 2,5 ч. Когда его выпустили, он отклонился от своего прежнего пути на такой же угол, на какой переместилось за это время солнце, и его лучи снова составили угол в 90° с направлением движения (По Brun, 1914.) | 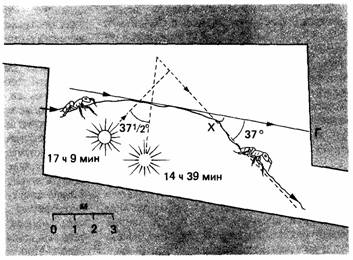
|
тация происходит под углом к направлению стимуляции. Примером служит реакция на «световой компас» у возвращающихся в муравейник муравьев. Эти животные отчасти ориентируются по положению солнца. Медленно меняя его видимое положение при помощи зеркала, можно заставить муравьев соответственно изменить свой путь (рис. 14.7). Раньше полагали (Brun, 1914), что если муравьев (Lasius niger), возвращающихся домой, посадить на несколько часов в темный ящик, то при освобождении они сохранят тот же угол движения по отношению к солнцу, что и до пребывания в ящике. Но затем стало очевидным (Jander, 1957), что муравьи делают поправку на перемещение солнца и по освобождении движутся в том же направлении. Такие компенсированные во времени компасные реакции продемонстрированы также у жука-навозника Geotrupes sylvaticus, прудовой водомерки Velia currens имедоносной пчелы Apis mellifera (см. обзор Saunders, 1976).
Тип ориентации, осуществляемый в этом случае, зависит как от природы внешних сигналов, так и от сенсорного оснащения организма. Животное, воспринимающее лишь силу стимула, ограничивается ее последовательными измерениями в разных пунктах. Если внешние сигналы по своей природе имеют направленность, единственный односторонне экранированный рецептор может предоставить дирекциональную информацию. Экранированный фоторецептор в этом отношении полезен, но экранированный хеморецептор никаких преимуществ не дает, потому что химические стимулы по природе своей не дирекциональны. При двух рецепторах одновременное сравнение может быть использовано для обнаружения градиентов (рис. 14.8). Если множество рецепторов расположено в форме растра (т.е. ряда или мозаики), возможны более сложные типы ориентации (рис. 14.8). Примерами растров служат глаза с хрусталиком у позвоночных и сложные глаза членистоногих (см. гл. 12). Пространственная ориентация часто достигается сочетанием разных методов. Например, самцы некоторых бабочек привлекаются самками с помощью испускаемого самкой и распространяющегося по воздуху феромона. Этот запах разносится ветром, поэтому летящий самец должен учитывать движение воздуха. Бабочки в полете обычно пользуются зрительными ориентирами, чтобы контролировать свое продвижение относительно почвы. На маршрут животного в воздухе влияет направление ветра, что и определяет его окончательный путь (рис. 14.9). Результаты опытов с бабочками показывают, что угол полета меняется с концентрацией запаха. В отсутствие запаха животное ле-
тает взад и вперед, не продвигаясь против ветра (т. е. под прямым углом к нему). Когда ветер приносит запах, угол полета увеличивается и животное движется зигзагами против ветра. Изменения направления связаны с границами запахового следа (рис. 14.9). При снижении концентрации запаха ниже определенного уровня, в частности у края его струи, животное начинает двигаться в противоположном направлении. Эти повороты не связаны с направлением ветра, а зависят от внутреннего эталона, или идиотетической информации. Таким образом в поисках полового партнера летящая бабочка использует сочетание зрительных, анемотаксических (связанных с ветром) и идеотетических ориентационных механизмов.
Дата добавления: 2016-06-18; просмотров: 2687;
Поиск по сайту
Узнать еще
- I. Ориентация формы в среде.
- А) пространственная структура.
- Виды дифференциации продукта. Горизонтальная (пространственная) продуктовая дифференциация. Модель линейного города (модель Хотеллинга). Модель кругового города (модель Салопа)
- Волновой пакет. Видность. Пространственная и временная часть фазы волны.
- И «спортивная ориентация»
- Конструкция формы, как ее пространственная организация.
- Координация и ориентация
- Миграции птиц как биологическое явление. Зависимость распределения птиц от распределения кормов. Ориентация и навигация.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
