Превращения азотсодержащих веществ
В природе происходит постоянный круговорот азота (рисунок 4.4), в котором участвуют растения, животные, микроорганизмы. Азот входит в состав белков и продуктов их разложения (пептиды, аминокислоты, пуриновые и пиримидиновые основания), а также в состав мочевины и мочевой кислоты, азотсодержащего полисахарида хитина, гумусовых кислот.
Аммонифицирующие микроорганизмы осуществляют разложение органических веществ, переводя азот органических соединений в минеральную форму (аммонийные соли, соли азотистой и азотной кислоты, молекулярный азот и аммиак), а также органические вещества своих клеток.
Нитрифицирующие бактерии способны превращать аммиак в азотистую кислоту, а затем в азотную. Затем в процессе денитрификации происходит выделение свободного азота в атмосферу. Зеленые растения при синтезе белков способны использовать только азот минеральных солей - нитратов и нитритов (в меньшей степени аммонийных солей), свободный азот воздуха и связанный в органических соединениях азот им недоступен.
Азот из атмосферного воздуха способны ассимилировать только специальные азотфиксирующие бактерии.
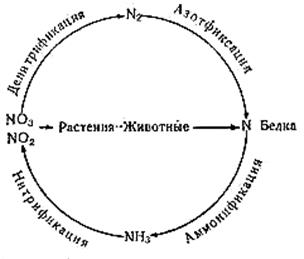
Рисунок 4.4 – Схема круговорота азота
Аммонификация
Аммонификация белка.Высокомолекулярные соединения, которыми являются белки, не могут непосредственно усваиваться микроорганизмами. Они должны быть расщеплены на составные части.
Процесс минерализации белковых веществ микроорганизмами с выделением аммиака или образованием аммонийных солей называется аммонификациейбелка, или гниением (рисунок 4.5), а микроорганизмы, вызывающие этот процесс – аммонифицирующим, или гнилостными микроорганизмами.
Аммонификация белков начинается с их гидролиза под действием протеолитических экзоферментов микроорганизмов. Происходит последовательное образование пептонов, пептидов и аминокислот.
Из полученных соединений наиболее доступными для микроорганизмов являются аминокислоты, которые растворимы в воде. В таком виде они поступают внутрь микробной клетки, где подвергаются воздействию эндоферментов, в частности дезаминированию под воздействием ферментов дезаминаз.
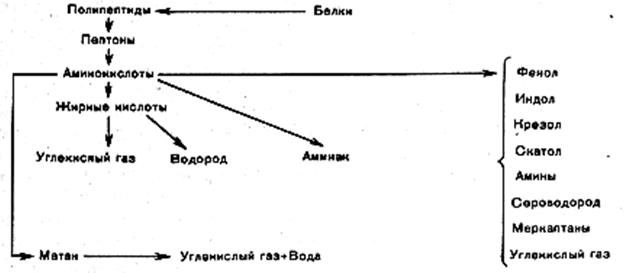
Рисунок 4.5 – Схема аммонификации белка
Существуют несколько типов дезаминирования (гидролитическое, окислительное, восстановительное и др.), которые приводят к образованию аммиака, оксикислот (гидрокислот), жирных кислот, кетокислот. Во всех случаях при любом типе дезаминирования среди конечных продуктов всегда обнаруживается аммиак:
+H2O
Гидролитическоедезаминирование: R-CHNH2-COOH → R-CHOH-COOH +NH3 (4.4)
гидрокислота
+H2
Восстановительноедезаминирование: R-CHNH2-COOH → R-CH2-COOH +NH3 (4.5)
жирная кислота
+O2
Окислительноедезаминирование: R-CHNH2-COOH → R-C=O-COOH +NH3 (4.6)
кетокислота
В аэробных условиях, кроме аммиака, образуется диоксид углерода, сероводород, в анаэробных - жирные и ароматические кислоты (бензойная и др.), спирты, индол, скатол, фенол, меркаптаны. В аэробных условиях происходит полное окисление жирных кислот с образованием СО2 и H2O.
Кроме дезаминирования внутри микробной клетки возможны и другие превращения аминокислот: декарбоксилирование, в результате которого появляются первичные амины (предшественники трупных ядов); переаминирование, приводящее к появлению новых аминокислот; синтез белка.
Декарбоксилирование происходит под воздействием эндоферментов декарбоксилаз с образованием СО2 и первичных (биогенных) аминов:
R-CHNH2-COOH → R- CH2 - NH2 + СО2 (4.7)
Сероводород образуется из аминокислот, содержащих серу:
+H2O
SH-CH2CH-NH2-COOH → CH2OH-CHOH-COOH + H2S + NH3 (4.8)
цистеин глицериновая кислота
В анаэробных условиях не происходит полного окисления жирных кислот, и они накапливаются в среде. Из ароматических аминокислот в этих условия образуются неприятно пахнущие ядовитые вещества – крезол, фенол, скатол, индол:
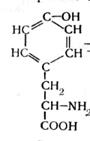

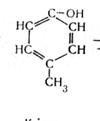

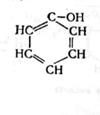
тирозин крезол фенол (4.9)
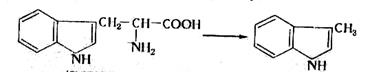
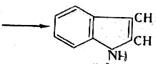 (4.10)
(4.10)
триптофан скатол индол
Из серусодержащих аминокислот образуются ядовитые меркаптаны:
SH-CH2CH-NH2-COOH → SH-CH2-CH3OH + СО2 + NH3 (4.11)
цистеин этил меркаптан
Биогенные амины являются токсичными соединения. Например, из лизина образуется кадаверин, из: орнитина – путресцин, аргинина – агматин, гистидина – гистамин.
NH2-(СH2)3-СH-NH2-COOH → NH2-(СH2)4-NH2 + СО2 (4.12)
лизин кадаверин
NH2-(СH2)4-СH-NH2-COOH → NH2-(СH2)5-NH2 + СО2 (4.13)
орнитин путресцин
Аммонифицирующие микроорганизмы. Аммонификацию белков вызывают микроорганизмы, обладающие протеолитическими ферментами (протеазами).
Аэробные аммонификаторы представлены в основном спорообразующими палочками рода Bacillus (B.mesentericus, B.subtilis, B.mycoides, B.megaterium и др.), а также неспорообразующими палочками рода Pseudomonas (например, P.fluorescens).
Факультативно-анаэробные аммонификаторы - это в основном представители семейства Enterobacteriaceae (Proteus vulgaris, Escherichia coli). Аммонифицирующие микроорганизмы встречаются среди актиномицетов, микроскопических грибов, (например, грибы родов Penicillium, Mucor, Apergillus, Trichoderma и др.).
Гнилостные анаэробные микроорганизмы – это спорообразующие бактерии рода Clostridium (C.perfringens, C.sporogenes, C.putrificum).
Аммонификация нуклеиновых кислот.Аммонификацию нуклеиновых кислот вызывают микроорганизмы, продуцирующие нуклеазы.Нуклеазы (внеклеточные ДНК-азы и РНК-азы) найдены у многих микроорганизмов – представителей родов Bacillus, Mycobacterium, Nocardia и др. (например, B.megatherium). При распаде нуклеиновых кислот и их производных (пуриновых и пиримидиновых оснований) тоже выделяется аммиак.
Аммонификация мочевины.Мочевина (карбамид) попадает во внешнюю среду в основном с мочой. Человек за сутки выделяет с мочой до 80 г мочевины. Азот в мочевине составляет 46%. За год люди и животные выделяют с мочой свыше 10 млн. т азота. Азот мочевины может усваиваться растениями только после ее разложения. Аммонификацию мочевины осуществляют микроорганизмы, продуцирующие фермент уреазу, под воздействием которого мочевина разлагается до углекислого аммония, который легко распадается на аммиак, углекислый газ и воду:
уреаза
(NH2)2CO + 2H2O → (NH4)2СО3 → 2 NH3 + СО2 + H2O (4.14)
Микроорганизмы, разлагающие мочевину, были открыты в 1862 г. Л.Пастером и названы уролитическими. Это аэробные бактерии, образующие споры. Среди них есть кокки (Micrococcus urea), сарцины (Planosarcina urea), бациллы (Bacillus probates, B.pasteurii). Благодаря образованию углекислого аммония при развитии уробактерий происходит подщелачивание среды (до рН 9…10). Уробактерии относятся к наиболее щелочелюбивым микроорганизмам.
Уробактерии обитают в почве, в рубце жвачных животных, в сточных водах. За счет разложения мочевины почва и водоемы обогащаются доступными для растений формами азота.
Аммонификация мочевой кислоты.Одним из конечных продуктов белкового обмена животных (птиц, пресмыкающихся, насекомых) является мочевая кислота. В помете птиц (гуано) она составляет 25%. В мочек млекопитающих концентрация мочевой кислоты незначительна. Разложение мочевой кислоты осуществляется рядом микроорганизмов (например, Clostridium acidiurici) с образованием мочевины и тартроновой кислоты:.
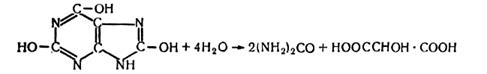 (4.15)
(4.15)
Аммонификация хитина. Хитин – очень стойкое соединение. Это сложный азотсодержащий полисахарид. Структурными элементами хитина являются остатки N-ацетилглюкозамина, соединенные 1,4-глико-зидными связями:
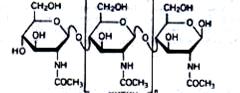 (4.16)
(4.16)
Хитин является основным компонентом клеточной стенки многих грибов (например, базидиомицетов, аскомицетов), а также панцирных покровов насекомых, ракообразных, некоторых беспозвоночных. После отмирания этих микроорганизмов хитин попадает в почву, водоемы, где разлагается микроорганизмами, продуцирующими хитиназы. Хитиназы широко распространены у актиномицетов, мукоровых грибов, представителей рода Bacterium (B. chitino-vorum, B.chitinophilum). В 1 г почвы содержится до 106 клеток микроорганизмов, использующих хитин. При разложении хитина образуется аммиак, глюкоза и уксусная кислота.
Аммонификация гумусовых веществ. Значительная часть связанного азота в почве содержится в форме перегнойных (гумусовых) веществ. Их разложение происходит медленно и осуществляется различными группами почвенных бактерий.
Нитрификация
Нитрификация – процесс окисления аммиака до азотистой, а затем азотной кислоты. Процесс в природе происходит в две фазы, каждая из которых осуществляется специфическими группами бактерий.
Первая фаза нитрификации– окисление аммонийных солей до солей азотистой кислоты (нитритов): 2NH3 + 3О2 → 2НNO2 + 2H2O + Эн.
Процесс осуществляют бактерии родов Nitrosomonas, Nitrosocystis, Nitrosospira, Nitrosovibrio, Nitrosolobus.
Вторая фаза нитрификации – окисление нитритов в нитраты: 2НNO2 + О2 → 2НNO3. Она осуществляется нитратными бактериями родов Nitrobacter, Nitrococcus, Nitrospira.
Нитрифицирующие бактерии открыл в 1889 г. С.Н. Виноградский. В настоящее время они объединены в семейство Nitrobacteriaceae. Это грамотрицательные микроорганизмы разнообразной формы, подвижные или неподвижные, аэробы или микроаэрофилы, развиваются в интервале рН 5…9,2.
Представителям некоторых родов свойственны обширные внутриклеточные разрастания ЦПМ. Особенностью бактерий рода Nitrosomonas является способность образовывать зооглейные скопления при повышенной концентрации нитритов. В этих скоплениях бактерии более устойчивы к неблагоприятным условиям.
Большинство этих бактерий являются облигатными автотрофами, которые при наличии органических веществ не размножаются. Лишь некоторые из них включают в состав своих клеток углерод отдельных органических соединений (глюкозы, аминокислот).
Основным источником углерода для нитрифицирующих бактерий является СО2 воздуха, ассимиляция которого осуществляется в цикле Кельвина, а необходимая для этого энергия поступает в результате окисления аммиака.
Энергетические процессы у нитрификаторов существенно преобладают над конструктивными. Они растут очень медленно, время генерации составляет около 24 ч.
Нитрификация происходит в почвах, озерах, морях, океанах. Она имеет как положительное значение, являясь важным этапом в круговороте азота в природе, так и отрицательное. Хотя нитраты хорошо усваиваются растениями и способствуют растворимости других необходимых растению элементов (например, фосфора и железа), они значительно легче и быстрее вымываются из почвы, чем аммонийные соли. Поступление азота с нитратами в воды морей и океанов составляет миллионы тонн в год. Поэтому интенсивная нитрификация может привести к обеднению почв азотом. С целью регуляции этого процесса вносят препараты, ингибирующие нитрификацию, например, нитропирин, пиридин и его производные. Эти препараты подавляют первую фазу автотрофной нитрификации, но не влияют на гетеротрофную нитрификацию.
Образовавшиеся при нитрификации нитраты подвергаются в почве дальнейшим превращениям: 1) они могут быть использованы высшими растениями; 2) могут вымываться в водоемы; 3) закрепляться в клетках микроорганизмов, которые используют нитраты в качестве источника азота; 4) восстанавливаться микроорганизмами до молекулярного азота и аммиака.
Денитрификация
Денитрификация – это восстановление, в результате которого азот переходит в менее окисленную форму, чем в нитратах. При денитрификации происходит восстановление нитратов до нитритов и далее до молекулярного азота: NO3- → NO2- → NО → N2O → N2
Этот перенос приводит к потере определенного количества почвенного азота и возврату его в атмосферу. В зависимости от того, какие микроорганизмы принимают участие6 в процессе денитрификации, различают прямую и косвенную денитрификацию.
Прямая денитрификация осуществляется непосредственно микроорганизмами, способными восстанавливать нитраты, используя их в качестве акцептора водорода при окислении ими органических веществ. Способностью к прямой денитрификации обладают многие почвенные микроорганизмы. Наиболее активными среди них являются бактерии рода Micrococcus (M. denitrificans), некоторые бациллы.
Микроорганизмы-денитрификаторы имеют две ферментные системы денитрификации: цитохромную (диссимиляционную) и флавопротеидную (ассимиляционную). Наличие этих систем обеспечивает два пути восстановления нитратов: ассимиляционный (неспецифический) и диссимиляционный (специфический).
Ассимиляционная денитрификация приводит к образованию аммиака, который ассимилируется организмом и включается в метаболизм. Нитраты здесь используются как источники азота.
Диссимиляционная денитрификация осуществляется микроорганизмами для получения энергии, ее конечным продуктом является молекулярный азот (рисунок 4.6).
.
4Н гидроксиламин Н2 Ассимиляционная
→ NН2OH → NH3 + Н2O денитрификация
Н2 Н2 ∕
NO3 → NO2 → NO
\ - Н2О Н2 Диссимиляционная
нитрат нитрит оксид → N2O → N2 + Н2O денитрификация
азота (II) оксид
азота (I)
Рисунок 4.6 – Схема денитрификации
Большинство денитрификаторов являются хемоорганотрофами. Использование в качестве конечного акцептора электронов нитратов позволяет им окислять органические субстраты до СО2 и воды.
Однако есть один вид денитрификаторов, который способен усваивать СО2 воздуха. Это Thiobacillus denitrificans. В отсутствие органических соединений он окисляет серу до серной кислоты за счет кислорода нитратов и полученную энергию использует для синтеза органических соединений из СО2 воздуха
Денитрифицирующие бактерии – факультативные анаэробы, переключающиеся на денитрификацию только в отсутствие кислорода. В аэробных условиях эти микроорганизмы окисляют органические вещества.
В клетках денитрификаторов есть ферментная система нитратаза, активизирующая кислород нитратов, и система, активизирующая молекулярный кислород. В анаэробных условиях функционирует первая ферментная система, в аэробных – вторая.
Косвенная денитрификация – чисто химический процесс, реакция восстановления, происходящая при взаимодействии нитратов с аминосоединениями, которые образуются в процессе жизнедеятельности различных микроорганизмов. В результате таких реакций тоже образуется молекулярный азот:
НNO2 + R-CHNH2COOH → R-CHOHCOOH + N2 + Н2O (4.17)
Микроорганизмам в этой реакции принадлежит косвенная роль, они образуют исходные продукты реакции – нитриты и аминосоединения.
Косвенная денитрификация имеет место только в кислой среде и значение ее невелико, так как окультуренные почвы не кислые. Более распространена в природе прямая денитрификация.
Денитрификация - широко распространенный процесс, в результате которого в атмосферу ежегодно поступает свыше 300млн. т азота. Особенно активно денитрификация протекает в ризосфере растений, где имеются органические вещества. Денитрификации способствует:
-слабая аэрация,
- высокая влажность,
- наличие достаточного количества органических соединений.
Денитрификацию не следует рассматривать как процесс, всегда приводящий к уменьшению азота в почве. Обычно в почве этот процесс до конца не доходит, так как растения усваивают образующиеся промежуточные продукты, возвращая их в круговорот. В настоящее время установлена даже положительная роль денитрификаторов в ризосфере растений, так как они способны кроме денитрификации одновременно осуществлять синтез веществ, стимулирующих рост растений.
Азотфиксация
Фиксация атмосферного азота в природе происходит физико-химическим и биологическим путем. Небиологический путь фиксации азота (грозовые разряды, работа двигателей внутреннего сгорания) дает всего 0,5% фиксированного азота. При электрических разрядах азот вступает в реакции с кислородом или водородом и попадает на землю в виде оксидов азота или аммиака.
Основной путь пополнения дефицита азота в почве – процесс биологической азотфиксации. Способность фиксировать молекулярный азот – уникальный процесс, присущий только прокариотам. За счет деятельности азотфиксаторов в почву ежегодно поступает 60…75% азота от общего его содержания в почве.
Способность микроорганизмов фиксировать азот была установлена С.Н. Виноградским и голландским ученым М. Бейеринком. Эта способность присуща многим систематическим группам бактерий (клостридии, сульфатредуцирующие бактерии, энтеробактерии, фотосинтезирующие спириллы, актиномицеты и многие другие группы прокариот). Существуют свободноживущие и симбиотические азотфиксаторы.
Свободноживущие азотфиксаторы. Представителем этих микроорганизмов является Clostridium pasteurianum, выделенный в 1893 г. С.Н. Виноградским. Это спорообразующие грамположительные анаэробные палочки, имеющие размеры 1,5х10 мкм. Они широко распространены в природе, являются возбудителями маслянокислого брожения.
Бактерии рода Azotobacter выделены в 1901 г. М. Бейеринком. Это аэробные бактерии. Молодые клетки представляют собой грамотрицательные полиморфные палочки, но в зрелом возрасте могут иметь форму диплококков, покрыты толстой слизистой капсулой. Молодые клетки подвижны.
При росте на безазотистых питательных средах азотобактеры образуют слизистые выпуклые колонии, окрашенные в темно-коричневый цвет (A.chroococcum), в желтый (A.vinelandii) и другие цвета.
Источником углерода и энергии для азотобактера служат углеводы, спирты, органические кислоты. Источниками азота, кроме молекулярного, могут быть соли аммония, нитраты, нитриты, аминокислоты, мочевина. При культивировании на азотсодержащих средах азотобактер утрачивает азотфиксирующую способность.
Аэробные азотфиксирующие бактерии рода Beijerinkia имеют морфологическое сходство с азотобактером, но отличаются медленным ростом м высокой кислотоустойчивостью (развиваются при рН 3,0). К свободноживущим азотфиксаторам относятся также фотосинтезирующие бактерии, некоторые микобактерии, актиномицеты, синезеленые водоросли.
Симбиотические азотфиксаторы. Это клубеньковые бактерии, впервые обнаруженные в 1866 г. М.С.Ворониным на корнях люпина и ольхи в клубеньках. Клубеньковые бактерии чаще всего обнаруживаются там, где растут бобовые растения, хотя могут встречаться в почве и в свободном состоянии.
Клубеньковые бактерии объединены в род Rhizobium (греч. rhizo – корень, bios – жизнь). Они проникают через корневые волоски в корневую систему растений и стимулируют деление тетраплоидных клеток корня, приводящее к образованию клубеньков. В клубеньках происходит интенсивное размножение бактерий. В молодых клубеньках большинство бактерий имеет форму палочек, с возрастом появляются клетки неправильной формы – бактероиды (L V X ), в которых происходит фиксация азота.
Клубеньковые бактерии характеризуются:
- специфичностью – способностью образовывать клубеньки только на корнях определенных видов бобовых растений;
- вирулентностью– способностью проникать через корневые волоски в корни растений и там образовывать клубеньки;
- активностью – интенсивностью фиксации азота.
Активность клубеньковых бактерий связана с наличием в них гемоглобина, который называется леггемоглобин. Это единственный представитель гемоглобинов, обнаруженных в растительном мире. Образование его возможно только в симбиозе бактерий с растениями.
Леггемоглобин выполняет функции регулятора парциального давления кислорода и транспорта электронов в азотфиксирующую систему.
Отношения между клубеньковыми бактериями и бобовыми растениями – пример мутуализма (симбиоза, при котором оба симбионта извлекают выгоду от совместного сожительства: растение получает азот, а бактерии используют корневые выделения ).
В настоящее время установлена способность ризобий осуществлять фиксацию атмосферного азота вне симбиоза с растениями. Доказана также способность к азотфиксации у представителей рола Escherichia, живущих в качестве комменсалов в пищеварительном тракте человека, найдены азотфиксаторы в кишечнике некоторых животных.
Дата добавления: 2017-11-21; просмотров: 7152;
Поиск по сайту
Узнать еще
- I.2.2 ТЕПЛОЁМКОСТЬ ВЕЩЕСТВА
- I.3.4 ВЗАИМНЫЕ ПРЕВРАЩЕНИЯ ЖИДКОСТЕЙ И ГАЗОВ
- II. Поведение веществ в магнитном поле.
- VI. Гемецеллюлозы и их превращения.
- VI. Пектиновые вещества и их роль в пищевой промышленности.
- А. Повторное применение лекарственных веществ
- Аварии с выбросом сильнодействующих ядовитых веществ
- Аварийно химически опасные вещества (АХОВ)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
