ОБЩАЯ ХАРАКТЕРИСТИКА
Грибы — обособленная группа гетеротрофных организмов, совмещающих признаки растений и животных.
С растениями их сближает наличие хорошо выраженной клеточной стенки (оболочки), неподвижность в вегетативном состоянии, размножение спорами, неограниченный рост, поглощение пищи путем абсорбции, т.е. всасывания всей поверхностью тела, а не заглатывания.
С животными их сближает гетеротрофность, наличие в клеточной стенке хитина, характерного для наружного скелета членистоногих, отсутствие в клетках хлоропластов и фотосин- тезирующих пигментов, накапливание гликогена как запасного вещества, образование и выделение продукта жизнедеятельности — мочевины. Эти анатомо-морфологические и физиоло- го-биохимические особенности грибов позволяют считать их одной из самых древних групп эукариотных организмов, не имеющих прямой эволюционной связи с растениями, как считалось ранее. Грибы и растения возникли независимо от разных форм водных протистов.
На основании этих данных с начала 70-х гг. XX в. грибы стали рассматривать в качестве самостоятельного царства органического мира наряду с царствами животных и растений (Тахтаджян, 1973; Whittaker, Margulis, 1978: Margulis, Schwartz, 1982 и др.) Хитридиомицеты, оомицеты, зигомицеты, аскоми- цеты, базидиомицеты получили статус самостоятельных отделов.
Грибы объединяют от 100 ООО до 250 ООО видов, широко распространенных на нашей планете. Они встречаются даже в песках пустынь, в морях и океанах, на скалах, высоко в горах и в полной темноте пещер.
Строение вегетативного тела грибов. Тело подавляющего большинства грибов представляет собой мицелий, состоящий из тонких бесцветных трубчатых нитей, или гиф, с верхушечным ростом и боковым ветвлением. У ряда грибов мицелий лишен перегородок (нечленистый, несептированный или ценоцитный) и представляет собой как бы одну гигантскую, часто сильно разветвленную клетку с большим количеством ядер. Ценоцитный мицелий известен у многих водных (сапро- легния и др.) и наземных грибов (мукор, ризопус и др.). Некоторые представители грибов образуют лишь зачатки мицелия в виде тонких безъядерных нитей — ризомицелий.
У большинства грибов гифы мицелия разделены поперечными перегородками, или септами, на отдельные клетки. Такой мицелий называют септированным или клеточным. Перегородки у грибов образуются путем инвагинации (впячи- вания) плазматической мембраны внутрь клетки, к ее центру (центрипетально), где всегда остается небольшое отверстие — пора. Поры обеспечивают тесное взаимодействие между клетками мицелия за счет взаимного перемещения питательных веществ и даже миграции органелл (ядра, митохондрии и т.д.).
Дрожжи, как и внутриклеточные грибы-паразиты, мицелия не имеют. Вегетативное тело дрожжей состоит из одиночных почкующихся или делящихся клеток, а у внутриклеточных паразитов — из голого протопласта без клеточной стенки. Такое вегетативное тело обычно недолговечно, через несколько дней оно превращается в зооспорангий с подвижными спорами, которые снова заражают подходящего хозяина.
С филогенетических позиций наиболее совершенной формой тела гриба является септированный мицелий, имеющий неограниченный рост за счет деления клеток и способный тем самым эффективно использовать субстрат. Кроме того, мицелий дифференцируется на две функционально различные части: субстратный, служащий для прикрепления к субстрату, поглощения и транспортировки воды и растворенных в ней питательных веществ, и воздушный, поднимающийся над субстратом и образующий органы размножения.
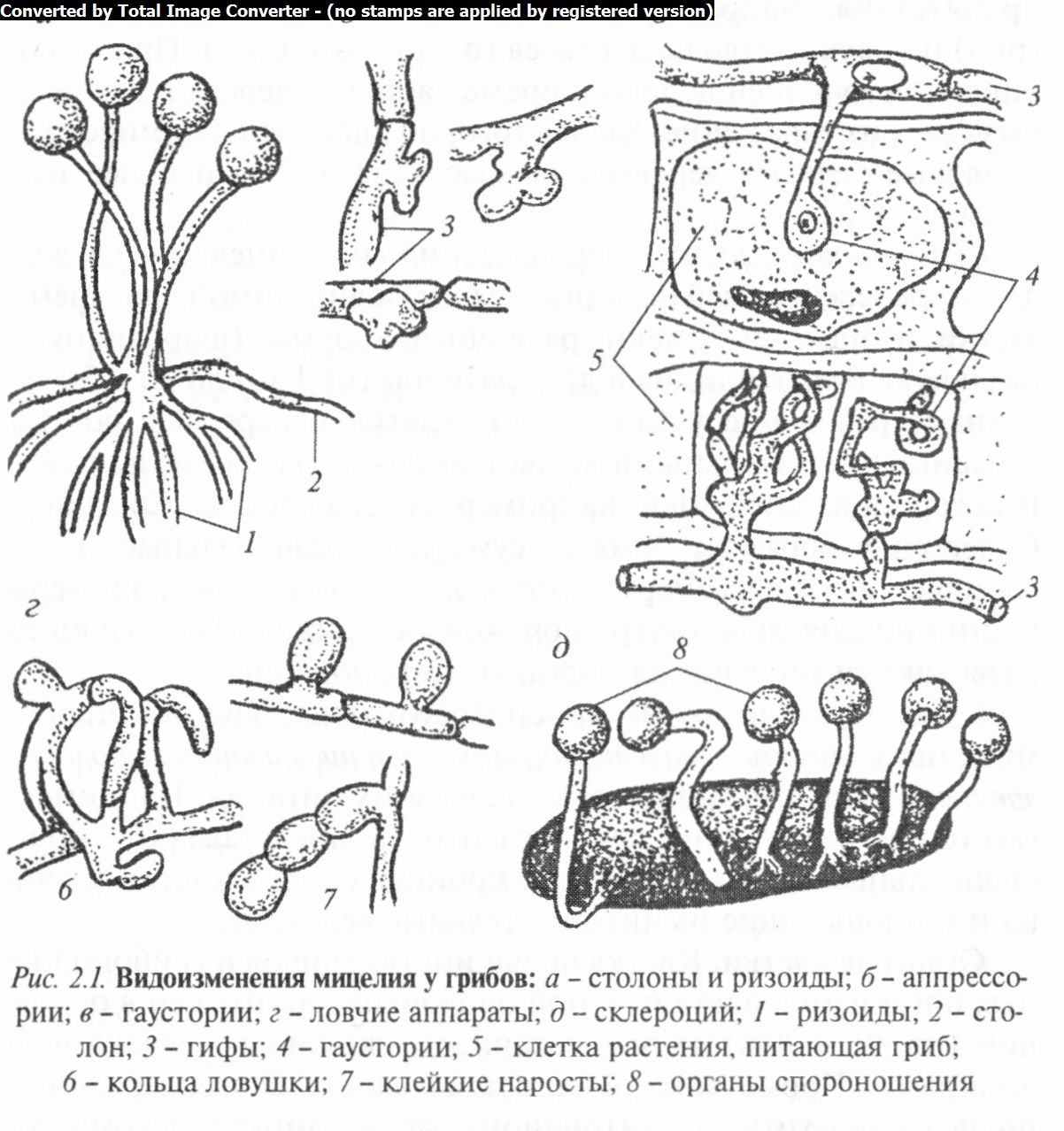
В процессе жизнедеятельности у грибов возникают многочисленные приспособления к различным условиям их обитания за счет видоизменения мицелия в ложную ткань — плектенхиму либо мицелиальные тяжи, ризоморфы, склероции и др. (рис. 2.1).
Плектенхима в отличие от настоящей ткани, образующейся при делении клеток во всех направлениях, формируется в результате переплетения и срастания нитей мицелия, клетки
•
которого делятся только в одном поперечном направлении. Из плектенхимы состоят плодовые тела шляпочных грибов.
Мицелиальные тяжи представляют собой плотно склеенные слизистым веществом пучки гиф толщиной до нескольких миллиметров. Они хорошо заметны у основания крупных плодовых тел шляпочных грибов, трутовиков, гастеромицетов в виде беловатых или окрашенных нитей. По ним протекают вода и питательные вещества.
Хорошо развитые и дифференцированные мицелиальные тяжи получили название ризоморф. Их наружные гифы имеют утолщенные темноокрашенные стенки и выполняют защитную функцию, а внутренние (тонкостенные и светлоокрашенные) — проводящую. Ризоморфы могут достигать нескольких метров в длину, позволяя грибам преодолевать значительные
препятствия, например участки каменных стен (домовой гриб) или сухие стволы деревьев (опенок осенний). При помощи ризоморф опенок легко перемещается от дерева к дереву и вызывает их заражение. Кроме того, он способен формировать плодовые тела на деревьях на высоте 2 — 3 м и более над землей.
Склероции — плотные переплетения гиф мицелия — служат для перенесения неблагоприятных условий зимой, во время засухи и т.д. Они имеют различные формы (шаровидную, овальную, в виде рожков и др.), размеры (от 1 мм до 20 — 30 см в диаметре) и массу (до 20 кг). Клетки склероциев богаты запасными питательными веществами — гликогеном, жирами. В склероциях спорыньи, например, содержится до 30% жира. Склероции образуют многие сумчатые, базидиальные и несовершенные грибы. Формируются они либо свободно на поверхности мицелия, либо внутри пораженного органа. Из склероциев развиваются мицелий или органы спороношения.
Существуют и другие приспособительные видоизменения мицелия у грибов — гаустории, аппрессории, ризоиды, столоны, ловушки и др., связанные со способом их питания. Например, гаустории характерны для паразитов и представляют собой специальные веточки мицелия, проникающие в клетки хозяина и поглощающие из них питательные вещества.
Строение клетки. Клетка большинства грибов и грибоподоб- ных организмов одета плотной оболочкой, состоящей в основном (на 80 — 90%) из полисахаридов, связанных с белками и липидами. Кроме того, в ее состав входят азотсодержащий полисахарид хитин (у хитридиомикот, зигомикот, аскомикот, базидиомикот и анаморфных грибов), полифосфаты, пигменты, меланины, глюканы и другие вещества. У оомикот клеточная стенка включает комплекс глюкан-целлюлозу (редко — хитин).
Протопласт клеток содержит одно или несколько ядер, митохондрии, эндоплазматическую сеть, рибосомы, лизосомы, вакуоли, а также специфические для них структуры в виде пузырьков — ломасомы, которые образуются между клеточной стенкой и плазматической мембраной. Функция ломасом еще окончательно не установлена. Комплекс Гольджи обнаружен у небольшого числа грибов (некоторые представители хитридиомикот). Он типичен для грибоподобных организмов
Запасными питательными веществами являются гликоген, дисахарид трегалоза, сахароспирты (маннит, сорбит, ксилит), волютин, масло. Крахмал в клетках не образуется.
Размножение грибов. Для грибов характерно бесполое (вегетативное и собственно бесполое) и половое размножение.
Вегетативное размножение может осуществляться фрагментами мицелия или в результате распадения гиф на отдельные клетки, каждая из которых дает начало новому организму. Клетки с тонкими оболочками, начинающими отчленяться с кончика гиф, называются артроспорами (оидиями), а с толстыми темноокрашенными оболочками — хламидоспорами. Они хорошо переносят неблагоприятные условия и прорастают чаще всего мицелием. Дрожжевые грибы, сумкоспоры у тафри- нальных грибов и базидиоспоры некоторых головневых способны размножаться почкованием.
Собственно бесполое размножение осуществляется посредством специализированных эндо- и экзогенных спор. Эндогенные споры (голые подвижные в воде клетки со жгутиками — зооспоры, неподвижные — спорангиоспоры) характерны для большинства оомицетов и хитридиомицетов. Они развиваются внутри специализированных клеток, отчлененных перегородкой от кончика гифы. Зооспоры образуются в зооспорангиях, а неподвижные многоядерные, одетые оболочкой спорангиоспоры, — в спорангиях.
Спорангии развиваются на спорангиеносцах, представляющих собой вертикально расположенную гифу, отходящую от мицелия. Такое расположение спорангиев облегчает распространение спор токами воздуха.
Экзогенные споры, или конидии, образуются открыто на концах особых специализированных выростов мицелия, называемых конидиеносцами. Формы конидиального спороношения разнообразны и характерны преимущественно для несовершенных, сумчатых и базидиальных грибов, а также для оомицетов и зигомицетов, приспособившихся к наземному существованию.
Эволюция органов бесполого размножения у грибов тесно связана со средой обитания. Наиболее примитивные зооспо- рангии присутствуют в основном у водных грибов (очень редко у наземных), в то время как спорангии и конидии характерны исключительно для наземных организмов.
Половое размножение у грибов особенно многообразно. У грибоподобных организмов (отдел Оомикота) и некоторых грибов (отдел Хитридиомикота) оно может проходить в форме холо-, изо-, гетеро- или оогамии. В последнем случае развиваются женские (оогонии) и мужские (антеридии) органы полово го размножения. В оогониях образуется одна или несколько яйцеклеток. Оплодотворение яйцеклеток происходит сперматозоидами или содержимым антеридия, которое переливается в оогоний через специализированные выросты — отроги. У зигомицетов половой процесс происходит путем слияния содержимого двух часто внешне не различимых клеток на концах гиф (зигогамия или гаметангиогамия).
Характерной особенностью полового процесса у оомикот, хитридиомикот и зигомикот является то, что зигота (циста, зигоспора) после обязательного периода покоя редукционно делится и прорастает с образованием органов бесполого размножения. Следовательно, у большинства примитивных грибов вся жизнь проходит в гаплоидной стадии.
У высших грибов при половом процессе происходит слияние содержимого антеридия и женского полового органа (архикарпа), не дифференцированных на гаметы (сумчатые грибы), или слияние содержимого двух вегетативных клеток мицелия — соматогамия (сумчатые и базидиальные грибы). В обоих случаях вначале сливаются цитоплазмы (плазмогамия), а мужские и женские ядра располагаются друг около друга, образуя дикарионы. Дикариогическая стадия в цикле развития сумчатых грибов представлена только аскогенными гифами и кратковременна, у базидиомикот она занимает большую часть жизненного цикла. Независимо от этого впоследствии происходит слияние ядер — кариогамия. Образующееся диплоидное ядро без периода покоя редукционно делится с образованием гаплоидных спор.
Половое спороношение у грибов бывает в виде сумкоспор, или аскоспор, и базидиоспор. Аскоспоры образуются эндогенно внутри специализированных клеток, называемых сумками или асками, базидиоспоры — экзогенно на базидиях. Сумки и базидии могут развиваться непосредственно на мицелии либо внутри или на поверхности плодовых тел.
У некоторых грибов половой процесс заменяется парасек- суалъным циклом, основными этапами которого являются гете- рокариоз, слияние разнокачественных гаплоидных ядер и последующая рекомбинация генетического материала при митозе.
На первом этапе клетки мицелия содержат генетически неоднородные ядра (гетерокариоз) за счет миграции ядер из гиф одного мицелия в гифы другого через анастомозы (цито- плазменные мостики), а также вследствие воздействия мута генных факторов. После слияния таких ядер образуется диплоидное гетерозиготное ядро. Затем происходит его мито- тическое деление, сопровождаемое перекомбинацией генетического материала (парасексуальный процесс) и восстановлением гаплоидности. Таким путем появляются формы с новыми свойствами.
Гетерокариоз и парасексуальный процесс характерны преимущественно для несовершенных грибов, лишенных настоящего полового процесса, а также имеет место у некоторых аско- и базидиомикот, обеспечивая их широкую изменчивость по морфолого-культуральным признакам и степени их вирулентности и агрессивности.
В циклах развития грибов наблюдается чередование гаплоидной и диплоидной ядерной фаз. При этом у одних групп доминируют гаплобионты, у других — диплобионты, у третьих - гаплоидная и диплоидная фазы имеют одинаковую продолжительность.
Экологические группы грибов. Грибы широко распространены в природе и встречаются на самых различных субстратах. В процессе длительной эволюции у них выработался комплекс приспособлений к различным пищевым связям и местообитанию.
Наиболее обширна группа почвенных грибов (около 70 ООО видов), которые благодаря наличию многочисленных ферментов участвуют в разложении (минерализации) органических веществ с образованием гумуса. Близко к ним примыкают грибы, разрушающие лесную подстилку: опавшие листья, хвою. Это подстилочные сапротрофы, включающие многочисленные плесневые и шляпочные грибы.
Среди почвенных грибов выделяют копрофилы (обитают на навозных кучах, местах скопления помета животных и т.д.); кератинофилы (приурочены к жизни на рогах, копытах и волосах животных); ксилофиты (разрушают как живую, так и мертвую древесину); хищные (способны жить как сапротрофы, питаются нематодами). Характерную группу составляют домовые грибы - разрушители деревянных построек.
Кроме почвенных в природе существуют водные грибы, среди которых можно выделить сапротрофы, живущие на растительных остатках, паразиты водных растений и животных, а также грибы, вызывающие обрастание деревянных частей судов, пристаней и др.
Многочисленной группой грибов являются паразиты водорослей, грибов, высших растений, животных и человека. Некоторые грибы утратили способность к самостоятельному существованию и приспособились к взаимовыгодному сожительству (мутуализму) с растениями. Особой формой симбиотиче- ской связи грибов являются лишайники и микориза. В последнем случае мицелий гриба оплетает корни растений и проникает только под эпиблему (эктотрофная микориза) или в клетки паренхимы корня, где может образовывать клубки (эндотрофная микориза). Микоризный гриб помогает растению усваивать труднодоступные вещества гумуса и поглощать воду, активизирует ферменты растения. От высшего растения гриб получает безазотистые соединения (углеводы) и корневые выделения, которые способствуют прорастанию спор. Микориза обнаружена у большинства высших растений (кроме водных).
( Роль грибов в природе и жизни человека. Грибы наряду с бактериями играют важную роль в общем круговороте веществ в природе. Разлагая органические вещества отмерших растений и животных, они делают их доступными для автотрофных организмов, участвуют в образовании плодородного слоя почвы — гумуса, выполняют большую санитарную работу по очищению среды.
Грибы широко используются в народном хозяйстве. Дрожжи применяют в хлебопекарной, пивоваренной, винодельной и спиртовой промышленности. Грибы используются для получения белка, ферментов, витаминов, антибиотиков, лимонной кислоты, ростовых веществ, а также препаратов для биологических методов борьбы с вредителями сельскохозяйственных растений. Многие шляпочные грибы употребляются как ценный пищевой продукт: в сухом веществе их плодового тела содержится в среднем 20 — 40% белка, 17 — 60 - углеводов, 1,5 — 10 — липидов и 6 — 25% минеральных элементов, органических кислот, витаминов (А, В j, В2, РР), смол и эфирных масел, придающих грибам своеобразный запах и вкус.
Паразитируя на растениях и животных, а также развиваясь сапротрофно на пищевых продуктах и изделиях из дерева, кожи, бумаги, пластмассы, металла, стекла, на произведениях искусства, грибы вызывают их порчу и приносят громадный ущерб народному хозяйству.
Дата добавления: 2021-03-18; просмотров: 1260;
Поиск по сайту
Узнать еще
- I. Общая характеристика категории состояния как часть речи
- II. Лексико-грамматические разряды имен числительных. Их характеристика.
- II. Лексико-грамматические разряды местоимений. Их общая характеристика
- IV. ОБЩАЯ ХАРАКТЕРИСТИКА ВИРУСОВ
- U - образная характеристика
- XXIII. ОБЩАЯ ХАРАКТЕРИСТИКА
- А. Общая характеристика
- Абсолютные показатели экономичности и энергетическая характеристика оборудования

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
