НАРУШЕНИЯ РЕГУЛЯЦИИ ДЫХАНИЯ
Под регуляцией дыхания понимается приспособление деятельности дыхательных механизмов к обеспечению газовых потребностей организма в условиях изменяющегося действия факторов внешней и внутренней среды. Например, при физической работе, метаболизм, а, следовательно, и потребление кислорода и выделение углекислоты возрастают многократно по сравнению с покоем. В самом деле, количество крови, переносящей кислород и углекислоту, выбрасываемой сердцем в одну минуту, может во время напряженной работы возрасти с 5 – 6 до 20 – 25 л (т.е. в 4 – 5 раз); объем же воздуха, вентилируемый легкими – с 6-8 до 100 – 150 литров, т.е. в 15 – 20 раз. При этом каждое состояние организма требует оптимальное количество кислорода, углекислоты и рН внутренней среды организма – т.е. такое их содержание, которое обеспечивает обмен веществ, в точности характерный для данного состояния. Главная структура, управляющая дыханием – дыхательный центр (ДЦ).
В функциональном отношении понятие дыхательного центра можно определить в узком и широком смысле. В узком смысле слова под ДЦ надо понимать весьма ограниченную нейронную структуру, которая определяет ритмическое дыхание и без которой дыхание невозможно. Такая нейронная организация расположена в продолговатом мозге на уровне задвижки.
В широком смысле слова под дыхательным центром понимают совокупность структур мозга, так или иначе участвующих в управлении дыханием и в наиболее совершенном приспособлении к изменяющимся дыхательным потребностям организма. Среди этих структур – все этажи ЦНС, начиная от коры больших полушарий и кончая мотонейронами спинного мозга. Почему правомерно такое неоднозначное понимание дыхательного центра? Чтобы разобраться в этом, нам необходимо сделать небольшой экскурс в историю развития представлений о структуре и функциях дыхательного центра.
Где находится дыхательный центр? Вопрос этот совсем не праздный. Несмотря на два столетия упорных поисков (пожалуй, они начались даже гораздо раньше – еще во 2 веке н.э., с К. Галена, который показал, что дыхание после отделения головного мозга от спинного у животного прекращается), местоположение его до сих пор окончательно не определено. Если рассматривать головной мозг «в профиль», заметно деление его на ствол и надстволовые отделы. Стволовая часть представляет собой продолжение спинного мозга. Она, в свою очередь, состоит из продолговатого мозга и моста. Именно здесь находится ряд важных центров. Еще в XVII в. ученые смогли убедиться, что разрушение продолговатого мозга ведет к прекращению дыхания. А в первой половине следующего столетия французские ученые Ц. Легаллуа и М. Флоуранс уточнили локализацию «жизненного узла». Оказалось, что достаточно буквально укола в определенную точку продолговатого мозга, чтобы дыхательные движения подопытного животного прервались. В опытах на животных также выяснилось, что эта небольшая (всего каких-нибудь 10 мм в поперечнике) область продолговатого мозга, если ее изолировать от других структур, обеспечивает лишь короткие, судорожные вдохи (гаспы). Чтобы дыхательные движения были правильными, регулярными («эйпное», что означает «хорошее дыхание»), необходимо участие ряда нейронных образований, в частности, расположенных выше, в варолиевом мосту (т.н. пневмотаксический центр).
Большой вклад в изучение дыхательного центра внесли крупные отечественные ученые – Н.А. Миславский, М.В. Сергиевский. Они пришли к мнению, что важнейшая часть дыхательного центра лежит у самой средней линии ретикулярной формации ствола мозга: если разрушить эту «медиальную» (т.е. срединную) зону дыхательного центра, дыхание прекращается. Далее, раздражая отдельные точки данной области, исследователи получили сокращение мышц-вдыхателей или выдыхателей.
Следовательно, здесь можно различить «инспираторный» и «экспираторный» отделы дыхательного центра. Особенно убедительно показал это известный авторитет в данном вопросе – американский нейрофизиолог Р.Ф. Питс.
Итак, к середине XX века сложилась довольно стройная и, казалось бы, полная картина устройства дыхательного центра: он расположен в медиальной зоне ретикулярной формации продолговатого мозга и состоит из двух групп нейронов, управляющих мышцами – вдыхателями и – выдыхателями.
Но наука не стоит на месте. Внедрение в исследовательскую практику современных электрофизиологических методов вызвало настоящий переворот в представлениях о локализации ДЦ. Оказалось, что в этом «дыхательном центре»... нет дыхательных нейронов! Зато они оказались в латеральной (боковой) зоне той же ретикулярной формации. Вводя микроэлектроды, соединенные с усилителем, ученые показали, что в т.н. дорсальном дыхательном ядре, оказались главным образом инспираторные нейроны. Их аксоны идут в шейный отдел спинного мозга, где импульсация переключается на двигательные нейроны, а те в свою очередь передают возбуждение на диафрагму. В другом, вентральном дыхательном ядре, находят как инспираторные, так и экспираторные нейроны. Их проводящие пути доходят до грудного отдела спинного мозга, где переключаются на мотонейроны межреберных мышц. А как же работают дыхательные нейроны?
Хочу сразу подчеркнуть, что это самый запутанный вопрос физиологии дыхания. До сих пор считалось (и так еще многие считают) что в дыхательном центре есть группа клеток – водителей ритма (как в сердце), способных возбуждаться самопроизвольно, под действием собственных обменных процессов. Следует заметить, что в очень сложно устроенном дыхательном центре теплокровных животных клетки-пейцмеккеры ищут уже которое десятилетие – и все безуспешно. А может их там вообще нет? Тогда кто же запускает этот нейронный маятник? В последнее время крупнейшие физиологи дыхания (Бредли, Ойлер, Черняк, Бреслав, Глебовский) склоняются к мысли, что ДЦ не обладает автоматизмом. Отдельные популяции нейронов каким-то образом возбуждают друг друга – это напоминает часовой механизм. Чтобы понять, как это происходит, стали разрабатывать модели ДЦ, точнее – дыхательного ритмогенеза, т.е. происхождения ритмической активности этого центра.
Рассмотрим эти модели. Самой распространенной из известных моделей является реципрокная. Суть ее состоит в том, что инспираторные нейроны, возбуждаясь, затормаживают экспираторные, а когда возбуждаются последние – тормозятся инспираторные. Однако когда математики стали строить модель этого реципрокного механизма, оказалось, что она не работает: колебания маятника в виде поочередного возбуждения и торможения инспираторных и экспираторных нейронов неизбежно должны затухать. Значит, дело не так просто.
Значительно успешнее были попытки построения моделей генераторов дыхательного ритма в виде замкнутых контуров, по которым возбуждение циркулирует по кольцу (стековые модели). Одной из них является модель московского физиолога В.А. Сафонова (1993) который предположил, что каждый цикл начинается с активации раннего инспираторного нейрона. Затем возбуждение переходит на поздний инспираторный ранний и поздний экспираторный нейроны, и, наконец, снова на ранний инспираторный, после чего все повторяется вновь. При этом, возбуждаясь, каждый нейрон затормаживает два предшествующих ему по фазе. В этой модели полные инспираторный и экспираторный нейроны служат «выходными»: через них циклический возбудительный процесс из микрокомплекса направляется по эфферентным нервным путям и реализуется в конечном итоге в сокращениях соответствующих дыхательных мышц. Но мы недаром сравнили работу дыхательного центра с часовым механизмом, куда входит, в частности, маятник. И, поскольку, в составе самих нейронов самовозбуждающегося водителя ритма найти не удается, неизбежно встает вопрос о «пружине» в этих часах, т.е. каких-то внешних источниках энергии. А такие источники действительно существуют. Это, прежде всего известная Вам ретикулярная формация, чьими полноправными членами и являются дыхательные нейроны. Польские исследователи В.Каржевский и Х.Громыш (1982) считают, что ДЦ может генерировать ритмическую активность только в том случае, если в ретикулярной формации оказывается «критическая масса» возбужденных нейронов. А это возможно лишь при условии поступления возбуждающих стимулов, как с периферии, так и от вышележащих центров. Так появились наиболее обоснованные на сегодня т.н. трансформаторные модели ДЦ. Познакомимся с наиболее известной из них – моделью американских авторов Бредли и Кларка (1987).
Под влиянием хеморецептивной стимуляции возбуждается определенная группа инспираторных нейронов (рис. 27). Это – т.н. центральная инспираторная активность (ЦИА). Чем сильнее афферентация – а эта импульсация зависит главным образом от напряжения СО2 в ликворе – тем быстрее происходит нарастание ЦИА, тем резче, происходит вдох. Но тем скорее этот вдох и прекратится.
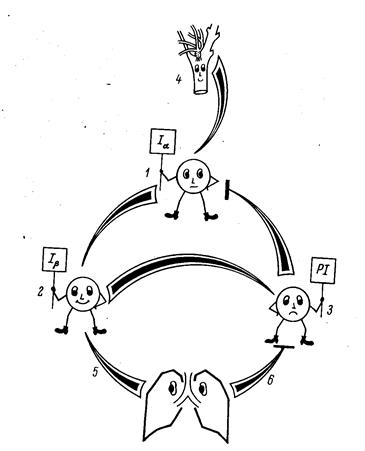
Рис. 27. Трансформаторная модель ритмогенеза
дыхательного центра Бредли и Кларка
Дело в том, что одновременно возбуждается другая группа нейронов, активность которой, достигнув определенной величины (порога) затормаживает, выключает ЦИА. Обрыву вдоха, переходу его в выдох способствуют также рецепторы растяжения легких. Вспомогательную роль в этом механизме играют влияния, поступающие сверху, прежде всего из пневмотаксического центра варолиева моста. Эти влияния способствуют более плавным переходам между инспираторной и экспираторной фазами и обратно. Эта модель иллюстрируется приведенной схемой. Здесь возбуждение инспираторных нейронов Ia (1) обеспечивает вдох. Одновременно активирующаяся другая популяция инспираторных нейронов (Ib) – 2 передает свое возбуждение, когда оно достигает пороговой величины постинспираторным нейронам (PI) – 3, которые, в свою очередь, затормаживают нейроны Ia, прекращая вдох и поддерживая некоторое время состояние выдоха. Активация нейронов Ia поддерживается и стимулами из хеморецептров (4). Раздражение рецепторов растяжения легких (5) способствует прерыванию вдоха, а ирритантных рецепторов (6) – прерыванию выдоха. Итак, мы можем констатировать: ДЦ, получая от хеморецептров информацию о химизме внутренней среды, преобразует ее в ритмический выходной сигнал (подобно трансформатору), обеспечивающий требующийся уровень вентиляции.
Мы подробно разобрали нейрофизиологические механизмы работы ДЦ для того, чтобы понять конкретные звенья нарушений регуляции дыхания. Расстройства регуляции в системе внешнего дыхания включают в себя:
1) центральные механизмы нарушений дыхательного ритмогенеза;
2) изменения в афферентном звене регуляции дыхательного центра (ДЦ);
3) нарушения проведения сигналов из ДЦ к респираторным мышцам.
Познакомимся с некоторыми терминами, отражающими изменения характера дыхательного рисунка:
1) гиперпное – увеличение глубины дыхательных движений, независимо от частоты;
2) тахипное – частое дыхание;
3) (полипное) – частое и глубокое дыхание;
4) брадипное – редкое дыхание;
5) апное – остановка дыхания;
6) диспное – одышка, некоординированное дыхание.
7) ортопное – выраженная одышка при застое в левом желудочке. В положении сидя ее переносить легче, застой уменьшается.
Расстройства центральной регуляции вентиляции легких возникают в основном вследствие нарушений функций дыхательного центра (ДЦ).
Исходя из изложенных выше представлений о структуре и функциях ДЦ, возможно выделить несколько механизмов расстройств регуляции дыхания, связанных с отклонениями в афферентной регуляции ДЦ и характеризующихся развитием гиповентиляционных и апноетических состояний.
а) Дефицит возбуждающей афферентации. Дефицит импульсов из хеморецепторов приводит к т.н. периодическим формам дыхания.

Рис. 28. Типы периодического дыхания
К ним относятся дыхание типа Чейн-Стокса – характеризуется постепенным нарастанием амплитуды дыхательных движений, сменяющегося их снижением и возникновением периода апное. Возникает при уменьшении чувствительности центрально-мозговых хеморецепторов к углекислому газу. Дыхание Биота – характеризуется чередованием серии частых и высоких дыхательных движений, сменяющихся периодами апное. Возникает при воспалительных и травматических поражениях мозга, опухолях задней черепной ямки. Кроме хеморецептивной стимуляции ранних инспираторных нейронов, большое значение имеют импульсы, поступающие из нейронов ретикулярной формации и парабрахиальных ядер пневмотаксического центра Люмсдена. Наблюдается при снижении тонической импульсации, например, нередко встречающегося в акушерской практике синдрома асфиксии новорожденных. Вследствие незрелости хеморецепторного аппарата ребенок, чаще недоношенный, может рождаться в состоянии асфиксии. Для активации ДЦ в таких случаях обычно используют дополнительные стимулирующие воздействия на кожные экстерорецепторы (похлопывание по ножкам и ягодицам ребенка, обрызгивание тела холодной водой и т.д.), ликвидируя тем самым дефицит возбуждающей афферентации через неспецифическую активацию ретикулярной формации. Уменьшение возбуждающей информации возникает при перерыве самого пневмотаксического центра (нижняя – средняя треть – возникает дыхание типа «айпнезиз»). Наконец, уменьшение притекающих импульсов наблюдается при ваготомии (двухсторонняя вагосимпатическая блокада) – при этом может возникать т.н. вагусная одышка (механизм компенсации – полисинаптические связи между ранними и поздними инспираторными нейронами в модели Бредли-Кларка).
Альвеолярная гиповентиляция и остановка дыхания, возникающие при угнетении ДЦ средствами для наркоза и обезболивания, также является следствием дефицита стимулирующих воздействий, необходимых для запуска генератора дыхательного ритмогенеза.
б) Избыток возбуждающей афферентации. «Перевозбуждение» ДЦ может характеризоваться развитием очень частого, но поверхностного дыхания. Альвеолярная гиповентиляция при тахипное (от греч. tachys-быстрый, скорый; pnoe – дыхание) является следствием увеличения функционального мертвого пространства. Причинами чрезмерной активации ДЦ могут быть стрессорные воздействия, приводящие к генерализованному возбуждению ЦНС, неврозы (например, приступы истерии), некоторые поражения структур среднего мозга, нарушения кровообращения, острое воспаление, механическая травма и др.
Избыток афферентации, возбуждающей ДЦ, может быть рефлекторного происхождения. Тахипноэ, сопровождаемое значительным уменьшением глубины дыхания, иногда возникает при раздражении брюшины, термических или болевых воздействиях на кожные покровы. Как отмечалось ранее, в механизме развития частого поверхностного дыхания (например, при крупозной пневмонии) существенное значение имеет раздражение ирритантных и юкстакапиллярных рецепторов легких.
Кашель может вызываться раздражением чувствительных окончаний плевры. Кашель состоит из короткого вдоха – затем немедленно смыкается голосовая щель и одновременно развивается экспираторное усилие дыхательных мышц. Резко повышается давление в дыхательных путях, голосовая щель немедленно раскрывается и воздух с большой силой выходит из дыхательных путей, вынося раздражающие слизью частицы. При чихании форсированный выдох происходит не через рот, а через нос.
в) Избыток тормозной афферентации. Этот механизм угнетения ритмической активности ДЦ включается, например, при раздражении слизистой оболочки верхних дыхательных путей в условиях развития острого респираторного заболевания воспалительного характера. Интенсивное раздражение слизистой оболочки полости носа и носоглотки химическими или механическими агентами может вызывать рефлекторную остановку дыхания на вдохе, повышение системного кровяного давления, брадикардию и даже кратковременное прекращение сердечных сокращений.
Этот тормозной тригеминовагусный рефлекс Кречмера возможно полностью предотвратить анестезией слизистой оболочки верхних дыхательных путей или в эксперименте – перерезкой ветвей тройничного нерва.
Дата добавления: 2016-11-29; просмотров: 3560;
Поиск по сайту
Узнать еще
- III. Механизмы регуляции количества ферментов
- А. Нарушения устной речи
- Административная ответственность за нарушения аграрного законодательства.
- Административная ответственность за экологические правонарушения
- Административные правонарушения в области воинского учета.
- Административные правонарушения, посягающие на права граждан.
- АКТИВНЫЕ ЭКТОПИЧЕСКИЕ НАРУШЕНИЯ РИТМА
- Анемии вследствие нарушения кровообразования

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
