Генетические основы дифференцировки
Яйцеклетка является уникальной клеткой, разные участки цитоплазмы которой имеют разный химический состав и различные потенции. В области анимального полюса цитоплазма обладает потенциями эктодермы, в области вегетативного полюса — энтодермы, а на экваторе — мезодермы (рис. 7.3).
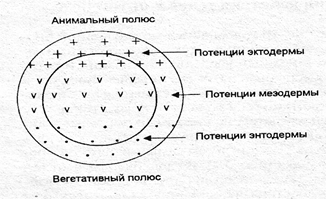
Рис. 7.3. Схема неоднородности цитоплазмы яйцеклетки (значки «+», «v», «•» обозначают разный химический состав цитоплазмы)
Приведем последовательность этапов дифференцировки:
1) первопричиной дифференцировки клеток является химическая разнородность цитоплазмы яйцеклетки (рис. 7.3), которая усиливается после оплодотворения (сегрегация);
2) химическая разнородность цитоплазмы яйцеклетки обеспечивает химическую разнородность цитоплазмы бластомеров (рис. 7.4); следовательно, разные бластомеры будут содержать разные индукторы;
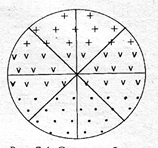
Рис. 7.4. Схема дробления зиготы
3) разные индукторы включают в работу различные транскриптоны;
4) синтезируются разные белки-ферменты;
5) разные белки-ферменты катализируют различные типы биохимических реакций;
6) в разных бластомерах идет синтез разных типо- и тканеспецифических белков, вследствие чего образуются различные типы клеток (проявление морфологической разнородности);
7) различные типы клеток образуют разные ткани;
8) из разных тканей формируются различные органы. Положение о том, что цитоплазма яйцеклетки содержит полный набор индукторов, способных включать в работу все необходимые блоки генов, подтверждают опыты Дж. Гердона (1964-1966). В яйцеклетку лягушки с удаленным ядром он пересаживал ядро соматической клетки (эпителия кишечника или кожи головастика). В большинстве случаев из такой яйцеклетки развивались нормальные головастики и даже взрослые лягушки (рис. 7.5). В 1996 г. в Шотландии аналогичными методами (клонирование) получены взрослые овцы. Эти опыты доказывают, что ядра соматических клеток содержат полную генетическую информацию о развитии целого организма, а цитоплазма яйцеклетки — полный набор индукторов для включения всех нужных блоков генов.
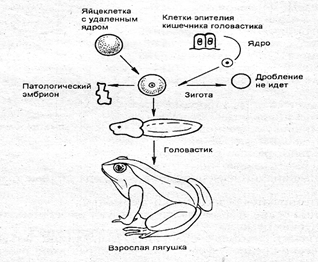
Рис. 7.5. Схема опытов Гердона
Опыты Гердона открыли возможность получения точных генокопий любого организма, в том числе и человека. Однако такие манипуляции с геномом человека чреваты серьезными морально-этическими последствиями. Для их проведения необходима соответствующая законодательная база. В последнее время большинство стран и многие ученые относятся к опытам по клонированию человека отрицательно. Условно гены можно разделить на 3 группы:
▪ функционирующие во всех клетках (например, гены, кодирующие ферменты энергетического обмена);
▪ функционирующие в клетках одной ткани (например, синтез миозина во всех клетках мышечной ткани);
▪ специфичные для одного типа клеток (например, гены гемоглобина в эритробластах).
Главный механизм дифференцировки — блокировка и деблокировка разных транскриптонов на каждом этапе онтогенеза (схема Георгиева). Так, при изучении гигантских хромосом из клеток слюнных желез насекомых удалось показать, что в отдельных местах хромосомы образуются вздутия — пуффы. Дезоксирибонуклеопротеидные нити в этих локусах деспирализованы и с них идет считывание информации. В зависимости от стадии развития пуффы появляются в разных участках нитей. Так, у личинки мухи дрозофилы в клетках слюнных желез одна из хромосом в конце третьей личиночной стадии имеет три характерных пуффа. Когда личинка превращается в предкуколку, они исчезают, но вместо них в другом локусе этой же хромосомы появляется характерный пуфф.
На ранних стадиях дробления бластомеры являются тотипотентными, т. е. каждый из них способен дать начало целому организму. В таких клетках может включаться в работу большинство блоков генов. Установлено, что у тритона тотипотентность сохраняется до стадии 16 бластомеров, у кроликов —до 4. Существование тотипотентности бластомеров у человека подтверждают случаи рождения нескольких монозиготных близнецов. Постепенно клетки становятся детерминированными, е. развитие их уже окончательно запрограммировано и они могут дать начало лишь клеткам определенного типа, например эпителиальным, нервным и др. В этих клетках могут работать только определенные блоки генов, а другие полностью заблокированы (выключены из работы).
Различают 2фазы дифференцировки: зависимую и независимую. В начале эмбриогенеза (до стадии ранней гаструлы) наблюдается зависимая дифференцировка, когда клетки еще относительно тотипотентны и их дифференцировка зависит от индукторов и соседних клеток. Например, клетки эктодермы спинной стороны зародыша хордовых дают начало нервной трубке, а брюшной — эпидермису кожи. Если взять клетки эктодермы с брюшной стороны зародыша и пересадить на спинную, они образуют нервную трубку.
На более поздних стадиях эмбриогенеза (стадия поздней гаструлы) клетки становятся детерминированными, их развитие предопределено и независимо от локализации они дифференцируются по намеченному плану — независимая дифференцировка. Например, если на стадии поздней гаструлы клетки верхней губы бластопора пересадить на нижнюю губу, то на брюшной стороне образуется нервная трубка.
Важную роль в развитии организма играет эмбриональная индукция — влияние группы клеток эмбриона на дифференцировку рядом расположенных клеток. Явление эмбриональной индукции было открыто Г. Шпеманом и Г. Мангольдом в 1924г. Первичный индуктор вызывает цепь последовательных вторичных индукций. Первичным индуктором являются клетки верхней губы бластопора, вызывающие дифференцировку клеток вентральной части энтодермы в хорду; хорда в свою очередь вызывает образование нервной трубки из дорсальной части эктодермы и пищеварительной трубки из вентральной части энтодермы (вторичная индукция). Так идет морфогенез — приобретение зародышем определенных морфологических структур.
В настоящее время считают, что эмбриональная индукция обусловлена выделением специфических химических веществ — индукторов, которые включают и выключают определенные блоки генов в близлежащих клетках.
В 30-е годы А.Г. Гурвичем и Н.К. Кольцовым была разработана концепция морфогенетических полей, которые интегрируют процессы развития в эмбриогенезе. Природа их точно не установлена. Она может быть разнообразной: электрической, гравитационной, термической и т. д. В это же время Ч. Чайлд ввел понятие градиента физиологической активности у зародыша, показав, что интенсивность обменных процессов постепенно падает от головного отдела к хвостовому. Это хорошо заметно у новорожденных: у них относительно большая и хорошо развитая голова, несколько хуже развиты верхние конечности и туловище и еще хуже — нижние конечности.
Дата добавления: 2016-11-26; просмотров: 3502;
Поиск по сайту
Узнать еще
- II. ОСНОВЫ МОЛЕКУЛЯРНОЙ ФИЗИКИ И ТЕРМОДИНАМИКИ
- V. ЛОГИЧЕСКИЕ ОСНОВЫ ВЫЧИСЛИТЕЛЬНОЙТЕХНИКИ
- XIV. ОСНОВЫ МАССОПЕРЕДАЧИ
- ІІ.5.2. Основы процесса фракталь-ного расширения квадрата
- Автогенетические процессы и половой отбор
- Анатомо - физиологические основы массажа
- Арифметические основы ЭВМ
- Атомное ядро и основы ядерной энергетики

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
