Биосинтез белка в клетке
Посредником в передаче генетической информации (порядок нуклеотидов) от ДНК к белку выступает информационная РНК. Она синтезируется в ядре на одной из цепей ДНК (кодирующей) по принципу комплементарности после разрыва водородных связей между двумя цепочками (фермент РНК-полимераза), причем считывание информации идет в одном направлении: 5΄→ 3΄. Процесс переписывания информации с ДНК на и-РНК называется транскрипцией. Синтезированная таким образом и-РНК (матричный синтез) выходит через поры ядра в цитоплазму и взаимодействует с малой субъединицей одной или нескольких рибосом, что приводит к сборке рибосомы (объединению большой и малой субъединиц). Рибосомы, объединенные одной молекулой и-РНК, называют полисомами. На каждой рибосоме полисомы синтезируются одинаковые молекулы белка.
Следующий этап биосинтеза белка — трансляция — перевод последовательности нуклеотидов в молекуле и-РНК в последовательность аминокислот в полипептидной цепочке. Транспортные РНК (т-РНК) "приносят" аминокислоты в рибосому. Молекула т-РНК по конфигурации похожа на лист клевера и имеет два активных центра. На одном конце молекулы расположен триплет свободных нуклеотидов, который называется антикодоном; он соответствует определенной аминокислоте. Так как многие аминокислоты кодируются несколькими триплетами, то число различных т-РНК значительно больше 20 (идентифицировано 60). Второй активный центр — противоположный антикодону участок, к которому прикрепляется аминокислота. На 5΄-конце этого центра молекулы т-РНК всегда находится гуанин, а на 3'-конце — триплет ЦЦА (рис. 3.15). Процесс узнавания т-РНК своей аминокислоты называется рекогницией. Каждая аминокислота присоединяется к одной из своих специфических т-РНК при участии особой формы фермента аминоацил-т-РНК-синтетазы и АТФ. В результате образуется комплекс аминокислоты с т-РНК — аминоацил-т-РНК, в котором энергия связи между концевым нуклеотидом А (в триплете ЦЦА) и аминокислотой достаточна для образования в дальнейшем пептидной связи. Аминокислоты транспортируются в большую субъединицу рибосом. В каждый данный момент внутри рибосомы находятся два кодона и-РНК: один — напротив аминоацильного центра, второй — напротив пептидильного центра.
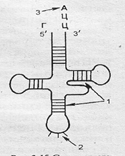
Рис. 3.15. Схема структуры молекулы т-РНК:
1- водородные связи; 2- антикодон; 3- место прикрепления аминокислоты
Если антикодон т-РНК и кодон и-РНК, находящийся напротив аминоацильного центра, являются комплементарными, то аминоацил-т-РНК присоединяется к кодону и-РНК, рибосома продвигается на один триплет и первая аминоацил-т-РНК оказывается в пепти-дильном центре. В аминоацильный центр поступает вторая т-РНК со своей аминокислотой. Между первой и второй аминокислотами устанавливается пептидная связь. Рибосома опять продвигается на один триплет; т-РНК первой аминокислоты отсоединяется от и-РНК и аминокислоты и уходит за следующей аминокислотой, а вторая т-РНК со своей аминокислотой попадает в пептидильный центр. В это время в аминоацильный центр поступает третья т-РНК с аминокислотой, и цикл повторяется. Таким образом, полипептидная молекула собирается в полном соответствии с информацией, записанной на и-РНК (рис. 3.16).
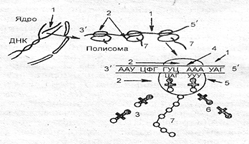
Рис. 3.16. Схема биосинтеза белка:
1- и-РНК; 2- субъединицы рибосомы; 3- т-РНК с аминокислотами,
4- колон и-РНК; 5- антикодонт-РНК; 6- т- РНК без аминокислот; 7- полипептид
В процессе трансляции выделяют три стадии: инициации, элонгации и терминации. Инициация (начало трансляции) заключается в связывании рибосомы с и-РНК, для чего в начале молекулы и-РНК имеется специальный инициирующий кодон (АУГ) и определенная последовательность нуклеотидов, которая отвечает за связь с рибосомой. Элонгация (непосредственная трансляция) включает реакции от образования первой пептидной связи до присоединения последней аминокислоты к молекуле полипептида. В это время рибосома перемещается от первого к последнему кодону на и-РНК. Терминация (конец трансляции) обусловлена наличием терминирующих кодонов (УАА, УАГ, УГА), которые прекращают синтез белка; происходит отделение рибосомы от и-РНК.
Регуляция синтеза белка у эукариот может осуществляться на уровне транскрипции и трансляции. Регуляторную функцию выполняют ядерные белки (гистоны). Их молекулы заряжены положительно и легко связываются с отрицательно заряженными фосфатами, влияя на транскрипцию определенных генов с помощью ДНК-зависимой РНК-полимеразы. Модификации гистонов (фосфорилирование, ацетилирование, метилирование) ослабляют их связь с ДНК и облегчают транскрипцию. Кислые негистоновые белки, связываясь с определенными участками ДНК, также облегчают транскрипцию. Регулируют транскрипцию и низкомолекулярные ядерные РНК, которые находятся в комплексе с белками и могут избирательно включать гены.
Усиливают синтез белка различные анаболические стероиды, инсулин, предшественники нуклеотидов и нуклеиновых кислот (инозин, оротат калия). Ингибиторами синтеза белка являются антибиотики (рифамицины, оливомицин), некоторые противоопухолевые препараты (винбластин, винкристин, 5-фторурацил), модифицированные азотистые основания и нуклеозиды.
Свойства генов
Гены характеризуются определенными свойствами: специфичностью, целостностью и дискретностью, стабильностью и лабильностью, плейотропией, экспрессивностью и пенетрантностью.
Специфичность гена заключается в том, что каждый структурный ген обладает только ему присущим порядком расположения нуклеотидов и детерминирует синтез определенного полипептида, р-РНК или т-РНК.
Целостность гена состоит в том, что при программировании синтеза полипептида он выступает как неделимая единица (цистрон), изменение порядка или количества нуклеотидов в которой приводит к перестройке структуры молекулы полипептида. Ген как функциональная единица неделим.
Дискретность гена определяется наличием в нем субъединиц (мутон, рекон). В настоящее время минимальной структурной субъединицей гена считают пару комплементарных нуклеотидов, а минимальной функциональной единицей — кодон.
Гены относительно стабильны и изменяются (мутируют) редко. Частота спонтанной мутации одного гена — примерно 10-5 на одно поколение.
Способность гена изменяться (мутировать) называется лабильностью.
Гены, как правило, обладают свойством плейотропности (множественности) действия, т.е. один ген отвечает за проявление нескольких признаков. Это, в частности, наблюдается при некоторых энзимопатиях, множественных врожденных пороках развития, например при синдроме Марфана.
Гены обладают также свойствами экспрессивности и пенетрантности (см. гл. 5).
Дата добавления: 2016-11-26; просмотров: 3504;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
