Клеточный и митотический циклы
Клеточный (жизненный) цикл — это период в жизнедеятельности клетки от момента ее появления до гибели или образования дочерних клеток. Митотический цикл — это период в жизнедеятельности клетки от момента ее образования и до разделения на дочерние клетки. Митотический цикл включает интерфазу и митоз (рис. 2.16).
Интерфаза состоит из трех периодов: пресинтетического (постмитотического) G1, синтетического S и постсинтетического (премитотического) — G2. Содержание генетической информации в клетке обозначают следующим образом: n — набор хромосом, хр — число хроматид в одной хромосоме, с — число молекул ДНК.
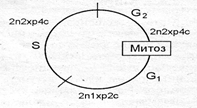
Рис. 2.16. Схема митотического цикла:
G1 - пресинтетический период; S- синтетический период; G 2 - постсинтетический период
Образовавшаяся после митоза клетка содержит диплоидный набор хромосом и соответственно удвоенное число молекул ДНК, каждая хромосома имеет одну хроматиду (2n1хр2с). Такая клетка вступает в пресинтетический период (G1) интерфазы, продолжительность которого колеблется от нескольких часов до нескольких месяцев и даже лет. В этот период клетка выполняет свои функции, увеличивается в размерах, в ней идет синтез белков и нуклеотидов, накапливается энергия в виде АТФ.
В синтетический период (S) происходит репликация молекул ДНК и ее содержание в клетке удваивается, т. е. каждая хроматида достраивает себе подобную, и генетическая информация к концу этого периода становится 2n1хр2с. Одновременно в клетке идут обменные процессы и она продолжает выполнять свои функции. Длительность этого периода 6-8 ч.
В постсинтетический период (G2) клетка готовится к митозу: накапливается энергия, синтезируются белки ахроматинового веретена.
Постепенно затухают все синтетические процессы, необходимые для репродукции органоидов, меняется вязкость ци-юилазмы и ядерно-цитоплазменное соотношение, прекращается выполнение клеткой основных функций. Содержание генетической информации не изменяется (2n2хр4с). Клетка вступает митоз.
Митоз
Митоз — это основной способ размножения соматических клеток. Главными причинами начала митоза являются: изменение ядерно-цитоплазменного соотношения (в разных клетках оно достигает 1/69—1/89);. появление "митогенетических лучей" — делящиеся клетки "заставляют" расположенные рядом клетки вступать в митоз; наличие "раневых гормонов" - поврежденные клетки выделяют особые вещества, вызывающие митоз неповрежденных клеток. Важная роль в регуляции деления клеток принадлежит белкам-циклинам, изменяющим продолжительность фазы G1 .
Непрерывный процесс митоза подразделяют на 4 стадии: профазу, метафазу, анафазу и телофазу, которым предшествует интерфаза (рис. 2.17).
В стадии профазы происходит увеличение объема ядра, спирализация хроматиновых нитей, расхождение центриолей к полюсам клетки и формирование веретена деления. К концу профазы фрагментируются ядрышки и ядерная оболочка, хромосомы выходят в цитоплазму и устремляются к центру клетки. В конце профазы к центромерам хромосом прикрепляются нити веретена деления. Содержание генетического материала при этом не изменяется (2n2хр4).

Рис. 2.17. Схема митоза в клетках корешка лука:
1- интерфаза; 2- профаза; 3- метафаза; 4- анафаза; 5- телофаза; 6- дочерние клетки
Метафаза — самая короткая фаза, когда хромосомы располагаются на экваторе клетки. В этой стадии достигается наибольшая спирализация хромосом и их удобнее всего изучать. Содержание генетического материала остается прежним.
В стадии анафазы происходит продольное разделение хроматид в области центромеры. Нити веретена деления сокращаются, и хроматиды (дочерние хромосомы) расходятся к полюсам клетки. Содержание генетической информации ставится 2n1хр2с у каждого полюса.
В стадии телофазы формируются ядра дочерних клеток: хромосомы деспирализуются, строятся ядерные оболочки, в ядре появляются ядрышки.
Митоз заканчивается цитокинезом — делением цитоплазмы материнской клетки. В конечном итоге образуются две дочерние клетки, каждая из которых имеет 2n хромосом, одну хроматиду в хромосоме и 2с наборов ДНК.
Основное значение митоза заключается в поддержании постоянства числа хромосом, обусловленном точным распределением генетической информации между дочерними клетками.
Разновидностями митоза являются эндомитоз, политения и мейоз. При эндомитозе происходит удвоение хромосом без деления ядра, что приводит к образованию полиплоидных клеток. При политении наблюдается многократное удвоение хроматид, но они не расходятся, и в результате образуются политенные (многохроматидные, гигантские) хромосомы, например в слюнных железах мухи дрозофилы.
Митоз — не единственный способ деления клеток. Эукариотические клетки могут размножаться и амитозом.
Амитоз — прямое деление клеток и ядер, находящихся в условиях физиологической и репаративной регенерации, либо опухолевых клеток. Типичный амитоз начинается с образования перетяжки ядра, затем цитоплазмы, и далее они делятся на две части. Установлено, что и при амитозе происходит равномерное распределение генетического материала между дочерними клетками.
Мейоз
Мейоз— это деление соматических клеток половых желез, в результате которого образуются половые клетки — гаметы. Мейотическое деление протекает в два этапа — мейоз-I и мейоз-П. Каждое мейотическое деление, так же как и митотическое, подразделяют на 4 фазы: профазу, метафазу, анафазу и телофазу (рис. 2.18).
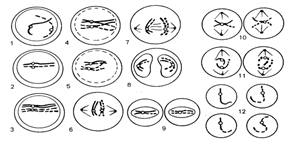
Рис. 2. I8. Схема мейоза (показана одна пара гомологичных хромосом):
Мейоз-I: 1- лептотена; 2- зиготена; 3- пахитена; 4- диплотена; 5- диакинез; 6- метафаза; 7- анафаза; 8- телофаза; 9- интеркинез; мейоз-П: 10- метафаза; 11-анафаза; 12- дочерние клетки
Наиболее сложной является профаза мейоза-I. Она подразделяется на 5 стадий: лептотена, зиготена, пахитена, диплотена и диакинез.
Хроматиновые нити спирализуются, вследствие чего они утолщаются и укорачиваются и на стадии лептотены становятся различимы под микроскопом. Во многих случаях заметны интенсивно окрашивающиеся участки — хромомеры, в которых хроматин сильно спирализован, и слабо окрашивающиеся, в которых хроматин слабо спирализован. Нитевидные гомологичные хромосомы начинают движение друг к другу центромерными участками. Содержание генетического материала составляет 2n2хр4с.
На стадии зиготены начинается конъюгация — попарное соединение гомологичных хромосом. Гомологичные хромосомы соприкасаются сначала в области центромер, а затем по всей длине. Их хромомеры точно совпадают. Содержание генетического материала не изменяется: 2n2хр4с.
На стадии пахитены гомологичные хромосомы тесно соприкасаются по всей длине, образуя биваленты. Бивалент — это пара гомологичных хромосом, каждая из которых состоит из двух хроматид, т. е. в биваленте содержится 4 хроматиды (отсюда другое название бивалентов — тетрады). Число бивалентов соответствует гаплоидному набору хромосом — 1 n. К концу этого периода начинают действовать силы отталкивания в области центромер, и становится заметным, что каждая хромосома состоит из 2 хроматид. Конъюгирующие хромосомы могут обмениваться участками хроматид —происходит кроссинговер (рис. 2.19). Содержание генетического материала не изменяется, однако его можно записать иначе — 1nбив4хр4с (1n бивалентов, каждый бивалент состоит из 4 хроматид и 4 наборов ДНК).
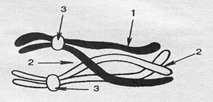
Рис. 2.19. Схема бивалента на стадии диплотены:
1 - хроматида; 2 - хиазмы; 3 – центромеры
На стадии диплотены между конъюгирующими гомологичными хромосомами продолжают действовать силы отталкивания, в результате чего хроматиды начинают расходиться, оставаясь соединенными в участках перекрестов — хиазм. Расхождение хроматид увеличивается, а хиазмы постепенно смещаются к их концам. Содержание генетического материала остается прежним (1nбив4хр4).
На стадии диакинеза завершается спирализация и укорочение хромосом (они окрашиваются равномерно). Биваленты, соединенные только своими концами, обособляются и располагаются по периферии ядра.
В течение профазы центриоли расходятся к полюсам клетки, а в конце профазы фрагментируются ядрышко и ядерная оболочка. Проконъюгировавшие хромосомы выходят в цитоплазму и движутся к экватору клетки. К центромерам хромосом прикрепляются нити ахроматинового веретена. Содержание генетического материала — 1nбив4хр4.
В метафазе мейоза-I в экваториальной плоскости клетки отчетливо видны биваленты, прикрепленные центромерами к нитям веретена деления. Содержание генетического материала остается прежним.
В анафазе мейоза-I гомологичные хромосомы, состоящие из двух хроматид, отходят к противоположным полюсам клетки. Расхождение хромосом носит случайный характер. Содержание генетического материала становится 1n2хр2су каждого полюса клетки, а в целом в клетке — 2(1n2хр2с). Телофаза мейоза-1не отличается от таковой митоза. В результате мейоза-I образуются две дочерние клетки, содержащие гаплоидный набор хромосом, но каждая хромосома имеет две хроматиды (1n2хр2с). Следовательно, в результате мейоза-I происходит редукция (уменьшение вдвое) числа хромосом, откуда и название этого деления — редукционное.
После окончания мейоза-I наступает короткий промежуток — интеркинез, в течение которого не происходит репликации ДНК и удвоения хроматид.
Мейоз-П — эквационное деление — протекает по типу обычного митоза. Профаза мейоза-П непродолжительная, так как хромосомы после телофазы мейоза-I остаются спирализованными. Изменений генетического материала не происходит (1n2хр2с). В метафазе мейоза-П хромосомы располагаются в экваториальной плоскости клетки. Содержание генетического материала — 1 1n2хр2с. В анафазу мейоза-П к полюсам отходят хроматиды (дочерние хромосомы) и содержание генетического материала становится 1nlxplc у каждого полюса клетки. В телофазе мейоза-П после цитокинеза образуются клетки с гаплоидным набором хромосом, содержащих одну хроматиду (1nlxplc).
Таким образом, в результате двух последовательных делений мейоза из одной диплоидной клетки образуются 4 гаплоидные.
Значение мейоза состоит в поддержании постоянства числа хромосом и рекомбинации генетического материала, обусловленной кроссинговером и случайным расхождением к полюсам гомологичных хромосом и хроматид.
При нарушении нормального течения митоза или мейоза наблюдаются различные хромосомные и геномные мутации — делеции, дупликации, полиплоидии, анеуплоидии и др.
Дата добавления: 2016-11-26; просмотров: 13055;
Поиск по сайту
Узнать еще
- II. Установление сегуната Токугава. Социально-экономическое развитие страны: циклы подъема и упадка
- Биогеохимические циклы (биогеохимические круговороты) -циклические процессы обмена веществ между различными компонентами биосферы, обусловленные жизнедеятельностью организмов.
- Вакуоли и клеточный сок
- Васкулиты крупных сосудов. Гигантоклеточный артериит
- Внутриклеточный поток энергии
- Гетероциклы с 2-мя атомами азота
- Гигантоклеточный артериит (височный артериит)
- Гуморальный иммунный ответ (В-клеточный).

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
