АНТИГЕНЫ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ.
В плазматических мембранах клеток разных тканей содержатся антигены главного комплекса гистосовместимости, которые играют важнейшую роль в иммунном ответе, иммунорегуляции, реакции отторжения трансплантата и других процессах. Для клинических и экспериментальных целей в качестве антигенов главного комплекса гистосовместимости определяют лейкоцитарные антигены. По своей химической природе эти антигены относятся к гликопротеинам клеточных мембран. По химической структуре и функциональному назначению ГКГ подразделяют на два класса.
1. ГКГ класса I состоят из двух полипептидных цепей с разной молекулярной массой: тяжелая ά-цепь (молекулярная масса 44000) нековалентно связана с легкой β-цепью (молекулярная масса 11600). Данные антигены содержатся в мембране почти всех ядросодержащих клеток. Они играют роль транспла-нтационных антигенов, варьирующих от человека к человеку и обеспечивающих реакцию отторжения трансплантата. Основная биологическая роль их состоит в том, что ГКГ -антигены класса I являются маркерами «своего», не подлежащего «атаке» Т-киллеров. При заражении клеток вирусами ГКГ -антигены класса I в комплексе с вирусными антигенами становятся своеобразными ориентирами для избирательного уничтожения зараженных клеток Т- киллерами.
2. ГКГ - антигены, принадлежащие к классу II. состоят из двух микроглобул и новых цепей примерно одной и той же молекулярной массы (34 000 и 28 000 соответственно), прикрепленных к поверхностной мембране макрофагов, Т- и В-лимфоцитов. Эти антигены участвуют в иммунорегуляции, служат для распознавания антигенных эпитопов Т-хелперами на мембране макрофагов и других клеток.
Генетический контроль ГКГ осуществляется генами, расположенными на хромосоме 6 в трех сублокусах: ГКГ - А, ГКГ -В, ГКГ - С.
Один человек не может иметь более 2 разных трансплантационных антигенов в одном сублокусе, т. е. не более 6 антигенов в трех сублокусах. ГКГ -сублокус находится в І -области хромосомы и содержит Іr гены (англ. immune response —иммунный ответ), контролирующие образование Іа- или ГКГ -DR-антигенов, принадлежащих к классу II.
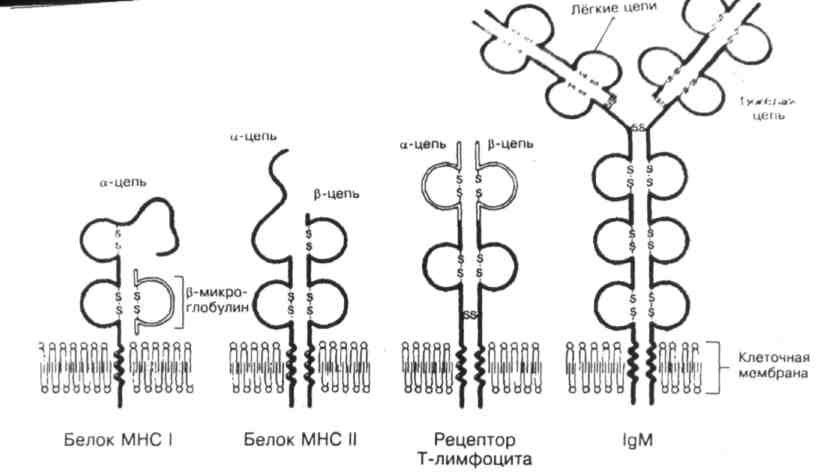
рис. 6. Структура белков семейства иммуноглобулинов ГКГ.
Молекула ГКГ I класса состоит из ά - цепи, внемембранная её часть связана с короткой цепью β - микроглобулина. Молекула ГКГ II класса состоит из двух СЕ: более длинной ά - цепи и β - цепи. Часть каждой цепи выступает на поверхности клеточной мембраны, цепь содержит трансмембранный участок и небольшой фрагмент в цитоплазме. Молекула рецептора Т-клеток состоит из двух цепей: ά и β. Каждая цепь представлена двумя внеклеточными Ig-подобными доменами, стабилизированными при помощи S- S связей, — вариабельным внеклеточным NH -концом и цитоплазматическим стабильным СООН - концом. SH-группа. присутствующая в цитоплазматическом фрагменте ά - цепи, может взаимодействовать с мембранными или цитоплазматическими белками. Мономер молекулы IgМ встраивается в плазматическую мембрану В-лимфоцитов, это рецептор Аг.
Главный комплекс гистосовместимости. Синтез главных Аг гистосовместимости (молекул ГКГ) контролирует комплекс генов ГКГ. Гены ГКГ расположены в коротком плече хромосомы 6, характеризуются выраженным полиморфизмом и имеют большое количество аллелей. Спектр молекул ГКГ уникален для каждого организма и определяет его биологическую индивидуальность.
Классификация. Молекулы, кодируемые генами ГКГ подразделяют на два класса: молекулы ГКГ I класса и молекулы ГКГ II класса.
Экспрессия. Молекулы ГКГ I и II классов — гликопротеины плазматической мембраны — экспрессированы на всех Аг-представляюших клетках и являются мишенями иммунного ответа при отторжении трансплантата.
а. Аг I класса представлены на поверхности практически всех клеток. Антигены ГКГ содержатся во многих органах и тканях. Самыми распростра-ненными из них являются антигены класса I. Они экспрессированы (представлены) почти на всех клетках, имеющих ядро; чаще на цитомем-бранах лимфоидных клеток, очень редко — на клетках мозга и скелетных мышц.
б. Аг II класса экспрессированы преимущественно на мембране иммунокомпетентных клеток, включая макрофаги, моноциты, Т- и В-лимфоциты. Антигены ГКГ класса II экспрессируются в основном на В-лимфоцитах и макрофагах, под воздействием γ-интерферона (цитокина) появляются на эндотелии капилляров и многих эпителиальных клетках.
Аномальная экспрессия ГКГ II. Неиммунокомпетентные эпителиальные клетки могут аномально экспрессировать белки ГКГ II. Белок ГКГ II, экспрессируемый на поверхности эпителиальных клеток, запускает каскад патологических реакций. Примеры:
(а) фолликулярные клетки щитовидной железы — раннее проявление ауто
иммунного заболевания щитовидной железы,
(б) эпителий жёлчных протоков при первичном жёлчном циррозе,
(в) β - клетки островков Лангергханса при сахарном диабете,
(г) эпителий слизистой оболочки кишечника при аутоиммунной затяжной диарее у детей.
Строение. Каждый Аг I и II классов содержит нековалентно связанные полипептидные цепи α и β. Цепи на 90% состоят из белка и на 10% из углеводов. Обе цепи кодируются близкорасположенными кластерами генов D региона. Большинство эпитопов (антигенных детерминант) расположено на β - цепи.
Функция. Молекулы ГКГ I и II класса контролируют иммунный ответ.
Антигены ГКГ выполняют в организме разнообразные функции, в частности определяют образование конвертаз и, следовательно, активацию комплеме-нта (антигены класса III); индуцируют процессы пролиферации и дифференциации клеток; служат объектом распознавания и отторжения трансплантатов; входят в состав рецепторов тимоцитов, гормональных рецепторов (антигены класса I). Наиболее значимым является то, что антигены МНС класса I и II определяют способность иммунной системы организма распознавать чужеродные антигены. Суть этого явления состоит в том, что микроорганизмы, продукты их распада или жизнедеятельности, другие нативные (неизмененные) антигены Т-лимфоцитами не распознаются. Все они вначале подвергаются процессингу (переработке) в эндосомах клеток, где происходят частичная денатурация и протеолиз, как правило, до пептидов. Такой низкомолекулярный процессированный антиген перемещается на поверхность клетки, связывается с находящимися здесь молекулами МНС класса I и II и становится доступным для восприятия Т-лимфоцитами. При этом надо подчеркнуть, что комплексы антигенов с молекулами МНС класса I распознаются цитотоксическими Т-лимфоцитами, которые осуществляют разрушение злокачественно перерожденных или инфицированных вирусом клеток, а комплексы антигенов с молекулами МНС класса II, образующиеся в основном на В-лимфоцитах и макрофагах, —Т-хелперами, которые как посредники передают сигнал и включают В- и Т-клетки в антителообразование или другие эффекторные процессы.
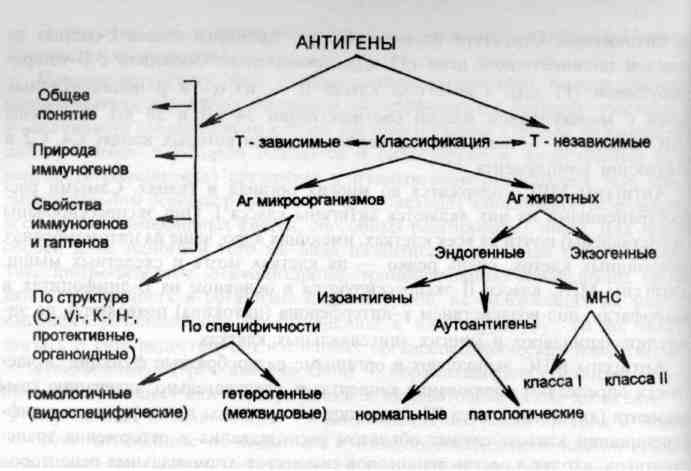
рис. 7. Виды антигенов.
Дата добавления: 2016-09-06; просмотров: 4805;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
