Краткая история становления и развития физиологии человека и животных
Наблюдения за жизнедеятельностью животных и человеческих организмов проводились с древних времен. За 14-15 веков до н.э. в Древнем Египте при изготовлении мумий люди хорошо знали внутренние органы человека. В гробнице врача фараона Унаса найдены изображения древних медицинских инструментов. В Древнем Китае только по пульсу удивительно тонко различали до 400 болезней. В IV- V веке до н. э. там уже существовало учение о функционально важных точках тела, которое в настоящее время явилось основой для современных методов диагностики и лечения. Древняя Индия прославилась своими особыми растительными рецептами, воздействием на организм упражнениями йоги и дыхательной гимнастики. В Древней Греции первые представления о функциях мозга и сердца высказывали в IV-V веке до н. э. Гиппократ (460-377 г. до н. э.) и Аристотель (384-322 до н. э.), а в Древнем Риме во 11 веке до н.э.— врач Клавдий Гален (201-131 г. до н. э.).
Как экспериментальная наука физиология возникла в XVII веке нашей эры, когда английский врач У. Гарвей открыл круги кровообращения. В этот же период французский ученый Р. Декарт ввел понятие рефлекс (отражение), описав путь внешней информации в мозг и обратный путь двигательного ответа. Работами гениального русского ученого М. В. Ломоносова и немецкого физика Г. Гельмгольца о трехкомпонентной природе цветного зрения, трактатом чеха Г. Прохазки о функциях нервной системы, наблюдениями итальянца Л. Гальвани о животном электричестве в нервах и мышцах отмечен ХVШ век. В ХІХ веке разработаны представления английского физиолога Ч. Шеррингтона об интегративных процессах в нервной системе, изложенные в его известной монографии в 1906 г. Проведены первые исследования утомления итальянцем А. Моссо. Обнаружил изменения постоянных потенциалов кожи при раздражениях у человека И.Р. Тарханов (феномен Тарханова).
В XIX веке работами основателя русской физиологии И.М. Сеченова (1829-1905) были заложены основы развития многих областей физиологии — изучение газов крови, процессов утомления и «активного отдыха», а главное — открытие в 1862 году торможения в центральной нервной системе и разработка физиологических основ психических процессов человека, показавших рефлекторную природу поведенческих реакций человека. Дальнейшая разработка идей И.М. Сеченова шла двумя путями. С одной стороны, изучение тонких механизмов возбуждения и торможения проводилось в Санкт-Петербургском Университете Н.Е. Введенским (1852-1922). Им создано представление о физиологической лабильности как скоростной характеристике возбуждения и учение о парабиозе как общей реакции нервно-мышечной ткани на раздражение. В дальнейшем это направление было продолжено его учеником А.А. Ухтомским (1875-1942), который, изучая процессы координации в нервной системе, открыл явление доминанты (господствующего очага возбуждения) и роль в этих процессах усвоения ритма раздражений. С другой стороны, в условиях хронического эксперимента на целостном организме, И.П. Павлов (1849-1936) впервые создал учение об условных рефлексах и разработал новую главу физиологии — физиологию высшей нервной деятельности. Кроме того, в 1904 г. за свои работы в области пищеварения И.П. Павлов, одним из первых русских ученых, был отмечен Нобелевской премией. Физиологические основы поведения человека, роль сочетанных рефлексов были разработаны В.М. Бехтеревым.
Крупный вклад в развитие физиологии внесли и другие выдающиеся отечественные физиологи: академик Л.А. Орбели, основавший эволюционную физиологию и адаптологию; академик К.М. Быков, изучавший условнорефлекторные влияния коры на внутренние органы; академик П.К.Анохин, создавший учение о функциональной системе; академик М.Н. Ливанов, основавший отечественную электроэнцефалографию; академик В.В. Ларин, разработавший космическую физиологию; Н.А. Бернштейн, основавший физиологию активности, и многие другие ученые-физиологи.
1.3 Общие закономерности физиологии и ее основные понятия
Живые организмы — это открытые системы, не замкнутые в себе, а неразрывно связанные с внешней средой. Они состоят из белков и нуклеиновых кислот и способны к авторегуляции и самовоспроизведению. К основным свойствам живого организма относятся: обмен веществ, раздражимость (возбудимость), подвижность, самовоспроизведение (размножение, наследственность), саморегуляция (поддержание гомеостаза, приспособляемость - адаптивность).
1.3.4 Основные функциональные характеристики возбудимых тканей
Общим свойством всех живых тканей является раздражимость, т.е. способность под влиянием внешних воздействий изменять обмен веществ и энергии. Среди всех живых тканей организма особо выделяют возбудимые ткани (нервную, мышечную и железистую), реакция которых на раздражение связана с возникновением специальных форм активности — электрических потенциалов и других явлений.
Основными функциональными характеристиками возбудимых тканей являются возбудимость и лабильность.
Возбудимость — свойство возбудимых тканей отвечать на раздражение специфическим процессом возбуждения. Этот процесс включает электрические, ионные, химические и тепловые изменения, а также специфические проявления возбудимости. В нервных клетках к таким проявлениям относятся импульсы возбуждения, в мышечных клетках — сокращение или напряжение, в железистых — выделение определенных веществ. Он представляет собой переход из состояния физиологического покоя в деятельное состояние. Для нервной и мышечной ткани характерна также способность передавать это активное состояние соседним участкам — т.е. проводимость.
Возбудимые ткани характеризуются двумя основными нервными процессами — возбуждением и торможением. Торможение — это активная задержка процесса возбуждения. Взаимодействие этих двух процессов обеспечивает координацию нервной деятельности в целостном организме.
Различают местное (или локальное) возбуждение и распространяющееся. Местное возбуждение представляет незначительные изменения в поверхностной мембране клеток, а распространяющееся возбуждение связано с передачей всего комплекса физиологических изменений (импульса возбуждения) вдоль нервной или мышечной ткани. Для измерения возбудимости пользуются определением порога, т.е. минимальной величины раздражения, при которой возникает распространяющееся возбуждение. Величина порога зависит от функционального состояния ткани и от особенностей раздражителя, которым может быть любое изменение внешней среды (электрическое, тепловое, механическое и пр.). Чем выше порог, тем ниже возбудимость, и наоборот. Возбудимость может повышаться в процессе оптимальной физической нагрузки и снижаться при утомлении.
Лабильность — скорость протекания процесса возбуждения в нервной и мышечной ткани. Понятие лабильности или функциональной подвижности предложено Н.Е. Введенским в 1892 г. В качестве одной из мер лабильности Н.Е. Введенский предложил максимальное количество волн возбуждения (электрических потенциалов действия), которое может воспроизводиться тканью в 1 с в соответствии с ритмом раздражения. Лабильность характеризует скоростные свойства ткани. Она повышается под влиянием раздражений, тренировки.
1.3.5 Нейрогуморальная регуляция функций
У простейших одноклеточных животных одна единственная клетка осуществляет разнообразные функции. Усложнение же деятельности организма в процессе эволюции привело к разделению функций различных клеток — их специализации. Для управления такими сложными многоклеточными системами уже было недостаточно древнего способа—переноса регулирующих жизнедеятельность веществ жидкими средами организма.
Регуляция различных функций у высокоорганизованных животных и человека осуществляется двумя путями: гуморальным (через кровь, лимфу и тканевую жидкость) и нервным.
Гуморальная регуляция функций действует сравнительно медленно и не может обеспечить срочных ответов организма (быстрых движений, мгновенной реакции на экстренные раздражители). В отличие от этого, нервная регуляция, осуществляемая нервной системой, обеспечивает быстрое и точное управление различными отделами целостного организма, доставку сообщений точному адресату. Оба эти механизма взаимосвязаны, однако ведущую роль в регуляции функций играет нервная система.
В регуляции функционального состояния органов и тканей принимают участие особые вещества — нейропептиды, выделяемые гипофизом и нервными клетками спинного и головного мозга. В настоящее время описано около 100 таких веществ, которые являются осколками белков и могут изменять функциональное состояние клеток, не возбуждая их. Они влияют на сон, процессы обучения и памяти, на мышечный тонус (в частности, на позную асимметрию), вызывают обездвижение или обширные судороги мышц, обладают обезболивающим эффектом.
1.3.6 Рефлекторный механизм деятельности нервной системы
В деятельности нервной системы основным является рефлекторный механизм. Рефлекс — это ответная реакция организма на внешнее раздражение, осуществляемая с участием нервной системы.Нервный путь рефлекса называется рефлекторной дугой. Обычно в состав рефлекторной дуги входят: воспринимающее образование — рецептор; чувствительный (афферентный) нейрон, связывающий рецептор с нервными центрами; промежуточные (вставочные) нейроны нервных центров; эфферентный (двигательный) нейрон, связывающий нервные центры с периферией; рабочий орган (эффектор), отвечающий на раздражение — мышца или железа. Наиболее простые рефлекторные дуги включают всего две нервные клетки, однако множество рефлекторных дуг в организме состоят из значительного количества разнообразных нейронов, расположенных в различных отделах центральной нервной системы. Выполняя ответные реакции, нервные центры посылают команды к рабочему органу (например, скелетной мышце) через эфферентные пути, выполняющих роль каналов прямой связи. В ходе рефлекторного ответа рецепторы, находящиеся в рабочем органе, и другие рецепторы тела посылают в центральную нервную систему информацию о результате действия. Афферентные пути этих сообщений — каналы обратной связи. Полученная информация используется нервными центрами для управления дальнейшими действиями, т. е. прекращением рефлекторной реакции, ее продолжением или изменением. Следовательно, основой рефлекторной деятельности являются не отдельные рефлекторные дуги, а замкнутые рефлекторные кольца, образованные прямыми и обратными связями нервных центров с периферией.
1.3.7 Гомеостаз
Внутренняя среда организма, в которой живут все его клетки, — это кровь, лимфа, межтканевая жидкость. Внутренняя среда характеризуется относительным постоянством — гомеостазом различных показателей, так как любые ее изменения приводят к нарушению функций клеток и тканей организма. К постоянным показателям гомеостаза относятся: температура внутренних отделов тела, сохраняемая в пределах 36-37° С; кислотно-основное равновесие крови, характеризуемое величиной рН = 7.4-7.35; осмотическое давление крови (7.6-7.8 атм.); концентрация гемоглобина в крови — 130-160 г. ּлֿ¹ и др.
Гомеостаз – это не статическое явление, а динамическое равновесие. Способность сохранять гомеостаз в условиях постоянного обмена веществ и значительных колебаний факторов внешней среды обеспечивается комплексом регуляторных функций организма. Эти регуляторные процессы поддержания динамического равновесия получили название гомеокинеза.
Степень сдвига показателей гомеостаза при существенных колебаниях условий внешней среды или при тяжелой работе у большинства людей очень невелика. Например, длительное изменение рН крови всего на 0,1 -0,2 может привести к смерти организма. При этом в общей популяции встречаются отдельные индивиды, которые могут переносить гораздо большие сдвиги показателей внутренней среды. У высококвалифицированных спортсменов-бегунов в результате большого поступления молочной кислоты из скелетных мышц в кровь во время бега на средние и длинные дистанции рН крови может снижаться до величин 7,0 и даже 6,9.
1.3.8 Возникновение возбуждения его проведение
1.3.8.1 Мембранные потенциалы. Мембрана клетки состоит из двойного слоя молекул липидов, между которыми свободно плавают глыбы белковых молекул. Некоторые из них пронизывают мембрану насквозь. В части таких белков имеются особые поры или ионные каналы, через которые могут проходить ионы, участвующие в образовании мембранных потенциалов (рис. I -А).
В возникновении и поддержании мембранного потенциала покоя основную роль играют два специальных белка. Один из них выполняет роль особого натрий-калиевого насоса, который за счет энергии АТФ активно перекачивает натрий из клетки наружу, а калий внутрь клетки. В результате концентрация ионов калия становится внутри клетки выше, чем в омывающей клетку жидкости, а ионов натрия — выше снаружи.
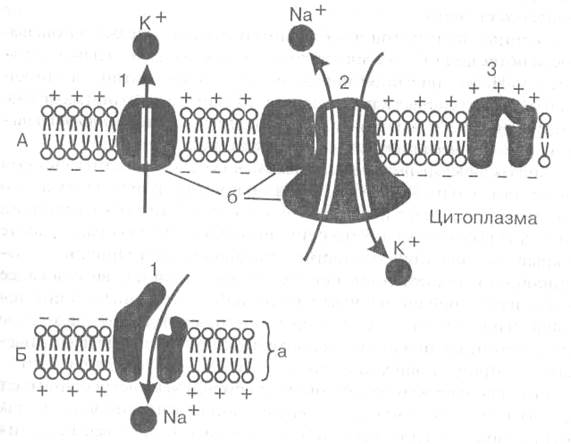
А — двойной слой липидов, б — белки мембраны.
А: каналы «утечки калия» (1), «натрий-калиевый насос» (2)
И закрытый в покое натриевый канал (3).
Б: открытый при возбуждении натриевый канал (1), вхождение ионов натрия в клетку и смена зарядов на наружной и внутренней стороне мембраны
Рисунок 1.1 – Мембрана возбудимых клеток в покое (А) и при возбуждении (Б) (По: Б.Альберте и др., 1986)
Второй белок служит каналом утечки калия, через который ионы калия в силу диффузии стремятся выйти из клетки, где они содержатся в избытке. Ионы калия, выходя из клетки, создают положительный заряд на наружной поверхности мембраны. В результате внутренняя поверхность мембраны оказывается заряженной отрицательно по отношению к наружной. Таким образом, мембрана в состоянии покоя поляризована, т. е. имеется определенная разность потенциалов по обе стороны мембраны, называемая потенциалом покоя. Она равна для нейрона примерно минус 70 м В, для мышечного волокна — минус 90 мВ. Измеряют мембранный потенциал покоя, вводя тонкий кончик микроэлектрода внутрь клетки, а второй электрод помещая в окружающую жидкость. В момент прокола мембраны и вхождения микроэлектрода внутрь клетки на экране осциллографа наблюдают смещение луча, пропорциональное величине потенциала покоя.
В основе возбуждения нервных и мышечных клеток лежит повышение проницаемости мембраны для ионов натрия — открывание натриевых каналов. Внешнее раздражение вызывает перемещение заряженных частиц внутри мембраны и уменьшение исходной разности потенциалов по обе стороны или деполяризацию мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению натрия внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные (локальные) изменения.
При увеличении раздражения изменения мембранного потенциала достигают порога возбудимости или критического уровня деполяризации — около 20 мВ, при этом величина потенциала покоя снижается примерно до минус 50 мВ. В результате открывается значительная часть натриевых каналов. Происходит лавинообразное вхождение ионов натрия внутрь клетки, вызывающее резкое изменение мембранного потенциала, которое регистрируется в виде потенциала действия. Внутренняя сторона мембраны в месте возбуждения оказывается заряженной положительно, а внешняя — отрицательно (рисунок 1.1-Б).
Весь этот процесс занимает 1-2 мс, после чего ворота натриевых каналов закрываются. К этому моменту достигает большой величины медленно нараставшая при возбуждении проницаемость для ионов калия. Выходящие из клетки ионы калия вызывают быстрое снижение потенциала действия. Однако окончательное восстановление исходного заряда продолжается еще некоторое время. В связи с этим в потенциале действия различают кратковременную высоковольтную часть — пик (или спайк) и длительные малые колебания — следовые потенциалы. Потенциалы действия мотонейронов имеют амплитуду пика около 100 мВ и длительность около 1,5 мс, в скелетных мышцах — амплитуда потенциала действия 120-130 мВ, а длительность 2-3 мс.
В процессе восстановления после потенциала действия работа натрий-калиевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, т. е. возвращение к исходной асимметрии их концентрации по обе стороны мембраны. На работу этого механизма тратится около 70% всей необходимой клетке энергии.
Возникновение возбуждения (потенциала действия) возможно лишь при сохранении достаточного количества ионов натрия в окружающей клетку среде. Большие потери натрия организмом (например, с потом при длительной мышечной работе в условиях высокой температуры воздуха) могут нарушить нормальную деятельность нервных и мышечных клеток и снизить работоспособность организма. В условиях кислородного голодания тканей (например, при наличии большого кислородного долга во время мышечной работы) процесс возбуждения также нарушается из-за поражения (инактивации) механизма вхождения в клетку ионов натрия, и клетка становится невозбудимой. На процесс инактивации натриевого механизма влияет концентрация ионов Са  в крови. При повышении содержания Са снижается клеточная возбудимость, а при дефиците Са возбудимость повышается, и появляются непроизвольные мышечные судороги.
в крови. При повышении содержания Са снижается клеточная возбудимость, а при дефиците Са возбудимость повышается, и появляются непроизвольные мышечные судороги.
1.3.8.2 Проведение возбуждения. Потенциалы действия (импульсы возбуждения) способны распространяться по нервным и мышечным волокнам (рисунок 1.2).
В нервном волокне потенциал действия является очень сильным раздражителем для соседних участков волокна. Амплитуда потенциала действия обычно в 5-6 раз превышает пороговую величину деполяризации. Это обеспечивает высокую скорость и надежность проведения.
Между зоной возбуждения (имеющей на поверхности волокна отрицательный заряд и на внутренней стороне мембраны — положительный) и соседним не возбужденным участком мембраны нервного волокна (с обратным соотношением зарядов) возникают электрические токи — местные токи. Развивается деполяризация соседнего участка, увеличивается его ионная проницаемость и появляется потенциал действия. В исходной зоне возбуждения при этом восстанавливается потенциал покоя. Затем возбуждением охватывается следующий участок мембраны и т. д. Так с помощью местных токов возбуждение распространяется на соседние участки нервного волокна, т.е. происходит проведение нервного импульса. По мере проведения амплитуда потенциала действия не уменьшается, т. е. возбуждение не затухает даже при большой длине нерва.
 |
Рисунок 1.2 – Схемы сенсорного и двигательного нейронов
В процессе эволюции с переходом от безмиелиновых нервных волокон к миелиновым значительно повысилась скорость проведения нервного импульса. Для безмиелиновых (безмякотных) волокон характерно непрерывное проведение возбуждения, которое охватывает последовательно каждый соседний участок нерва. Миелиновые (мякотные) нервы почти полностью покрыты изолирующей миелиновой оболочкой. Ионные токи в них могут проходить только в оголенных участках мембраны — перехватах Ранвье, лишенныхэтой оболочки. При проведении нервного им пульса возбуждение перескакивает от одного перехвата к другому и может охватывать сразу несколько перехватов. При этом возрастает не только скорость, но и экономичность проведения. Возбуждение захватывает не всю поверхность мембраны волокна, а лишь небольшую ее часть. Значит, меньше энергии тратится на активный транспорт ионов через мембрану при возбуждении и в процессе восстановления.
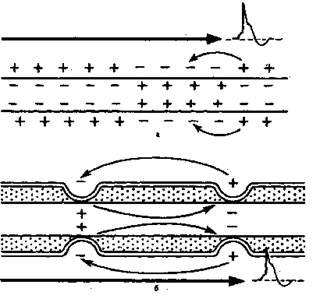 Скорость проведения в разных волокнах различна. Более толстые нервные волокна проводят возбуждение с большей скоростью: у них расстояния между перехватами Ранвье больше и длиннее скачки. Наибольшую скорость проведения имеют двигательные и проприоцептивные афферентные нервные волокна — до 100 м/с. В тонких симпатических нервных волокнах (особенно в немиелинизированных волокнах) скорость проведения мала — порядка 0.5 — 15 м/с (рисунок 1.3).
Скорость проведения в разных волокнах различна. Более толстые нервные волокна проводят возбуждение с большей скоростью: у них расстояния между перехватами Ранвье больше и длиннее скачки. Наибольшую скорость проведения имеют двигательные и проприоцептивные афферентные нервные волокна — до 100 м/с. В тонких симпатических нервных волокнах (особенно в немиелинизированных волокнах) скорость проведения мала — порядка 0.5 — 15 м/с (рисунок 1.3).
Рисунок 1.3 – Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
Во время развития потенциала действия мембрана полностью теряет возбудимость. Это состояние называют полной невозбудимостью, или абсолютной рефрактерностью. За абсолютной следует относительная рефрактерность, когда потенциал действия может возникать лишь при очень сильном раздражении. Постепенно возбудимость восстанавливается до исходного уровня.
Дата добавления: 2016-09-06; просмотров: 4950;
Поиск по сайту
Узнать еще
- Filariidae:. Onchocerca volvulus Систематика, морфология, цикл развития, патогенное действие, диагностика и профилактика онхоцеркоза и лоаоза.
- I. История открытия и методы исследования вирусов
- II. Завоевание Китая маньчжурами. Экономическое положение страны в XVII – начале XIX вв.: аграрная политика Цинской династии, особенности развития городского ремесла
- II. Особенности развития турецкой буржуазии. Становление младотурецкого движения
- III Всебелорусское собрание. Программа социально-экономического развития Республики Беларусь на 2006 – 2010 гг.
- IV. Внешние условия развития отрасли информационных технологий
- VI.3. Краткая характеристика классов месторождений
- VII. Пути развития национального экологического законодательства с учётом международных обязательств РК

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
