Ультраструктурная организация клеточной мембраны
Толщина мембраны составляет около 10 нм. Мембрана состоит главным образом из липидов и белков с примесью углеводов. Липиды представлены фосфолипидами, которые имеют полярные «головки» и неполярные «хвосты», т.е. на одном конце молекулы имеются заряженные ионные группы, а другой конец является электронейтральным. Полярные головки молекул стремятся контактировать с водой, а неполярные хвостовые части избегают таких контактов и притягиваются друг к другу. В результате образуются плёнки, состоящие из двух слоёв липидных молекул. Полярные головки обращены к наружной и внутренней среде клетки, где много воды, а неполярные «хвосты» расположены между ними, в середине мембраны.
Транспорт ионов через мембрану может осуществляться двумя путями: через ионные каналы, и с помощью молекул-переносчиков. Через ионные каналы перенос ионов осуществляется пассивно, так как сама клетка не затрачивает для этого свою энергию. Перемещение ионов происходит за счет осмотических сил, обусловленных разностью концентрации иона по обе стороны мембраны, по градиенту (разнице) концентрации данного иона – из среды с большей концентрацией в среду с меньшей концентрацией.
Активный транспорт ионов через мембрану происходит с участием переносчиков, или ионных насосов. В этом виде транспорта перенос ионов происходит против концентрационного градиента, то есть ионы переносятся в область более высокой их концентрации, и для этого клетка затрачивает значительную энергию, используя для этих целей АТФ.
Ионные каналы представляют собой тончайшие поры, образованные молекулами интегральных белков. Каналы пронизывают мембрану и соединяют наружную и внутреннюю среду клетки. Имеются разные каналы для ионов натрия, калия, кальция, хлора. Они отличаются диаметром и наличием заряженных структур в канале, дифференцирующих анионы и катионы. Каналы могут быть в трех состояниях: открытом (активированном), закрытом и инактивированном (неспособным открыться).
Бóльшая часть калиевых каналов практически всегда открыта. Натриевые и кальциевые каналы открываются только при действии на них раздражителей – либо электрического тока, либо некоторых химических веществ. Такие каналы соответственно называются электрозависимыми или хемозависимыми. В открытом (активированном) состоянии натриевые или кальциевые каналы находятся в течение очень короткого времени, измеряемого сотыми или тысячными долями секунды. Среднее время нахождение канала в открытом состоянии составляет менее 1 мс.
Ионные насосы - это полуинтегральные белковые молекулы, они располагаются либо на наружном, либо на внутреннем слое мембраны. На поверхности молекулы имеется группа атомов, присоединяющих к себе какие-либо ионы. Быстро перемещаясь на другую сторону мембраны, белковые молекулы переносят присоединившиеся ионы и освобождаются от них на другой стороне мембраны. Такой транспорт требует энергии и называется активным транспортом, или «насосом» с его помощью осуществляется перенос веществ (ионов) в среду с более высокой их концентрацией, то-есть против концентрационного градиента. Известны молекулы-переносчики для протонов, иода, кальция, натрия и калия.
Благодаря ионным насосам поддерживается ионный гомеостаз цитоплазмы. Необходимо помнить, что состав ионов в цитоплазме отличается от ионного состава среды, окружающей клетку – внутри клетки в несколько раз больше концентрация калия, чем вне ее, а снаружи – больше концентрация кальция и натрия, чем внутри клетки. Такое неравновесное содержание ионов по обе стороны мембраны и поддерживается работой переносчиков, или насосов. Так, кальциевый насос выкачивает из клетки излишний кальций, а калий-натриевый насос выкачивает из клетки ионы натрия в обмен на ионы калия. Работа ионных каналов происходит непрерывно, но во время возбуждения она увеличивается.
Потенциал покоя
Потенциал покоя возникает вследствие перемещения ионов калия из клетки во внешнюю (по отношению к клетке) среду. Это объясняется тем, что в состоянии физиологического покоя калиевые каналы в мембране открыты, а концентрация калия внутри клетки больше, чем снаружи. Положительно заряженные катионы калия, выходя через открытые калиевые каналы, и придают наружной поверхности мембраны положительный заряд. Внутренняя поверхность мембраны оказывается электроотрицательной по отношению к наружной. Величина потенциала покоя – 60 - 90 мВ.
Потенциал покоя может сохраняться на мембране весьма продолжительное время, пока ткань не будет переведена в состояние возбуждения.
Значение потенциала покоя: транспорт веществ через мембрану, регуляция межклеточных взаимодействий. Например, электрическое поле, имеющееся на мембране, втягивает внутрь клетки одни вещества и не пропускают другие; одноименно заряженные эпителиоциты и форменные элементы крови не прикасаются к стенкам кровеносных сосудов, а клетки крови не склеиваются.
Хотя большинство натриевых каналов в состоянии покоя закрыто, небольшое количество катионов натрия все же проникает внутрь клетки с помощью переносчиков. Так, например, некоторые вещества (глюкоза, аминокислоты) могут попадать в клетку только в соединении с ионами натрия. Внутри клетки проникшие вещества отделяются от переносчика, и содержание натрия в цитоплазме увеличивается.
Поскольку пассивный выход калия из клетки и одновременно небольшой вход натрия осуществляются непрерывно, то и ионные насосы должны работать постоянно, сохраняя разность концентрации ионов в цитоплазме и окружающей среде. Поэтому одновременно с пассивным током калия из клетки работает и система переносчиков, возвращающих калий обратно в клетку, и выкачивающих из клетки ионы натрия. Такие переносчики называются калий-натриевыми насосами. Благодаря их работе сохраняется высокая концентрация калия внутри клетки, и низкая - натрия.
Таким образом, в состоянии покоя пассивный ток калия из клетки создает разность потенциалов на мембране (снаружи плюс, внутри минус), а активный транспорт с помощью калий-натриевого насоса поддерживает разность концентрации этих ионов. Оба процесса происходят одновременно и непрерывно.
Потенциал действия
При действии на ткань раздражителя открываются натриевые каналы. В этом и заключается суть действия раздражителя. Поскольку концентрация ионов натрия внутри клетки меньше, чем во внеклеточной среде, через открывшиеся натриевые каналы катионы натрия начинают пассивно проникать внутрь клетки.
Мембранный потенциал сначала уменьшается и исчезает (деполяризация мембраны), а дальнейший ток натрия внутрь приводит к тому, что внутренняя поверхность мембраны становится электроположительной, а наружная – электроотрицательной. Величина потенциала действия – 110 - 130 мВ.
На схеме обозначены 5 фаз потенциала действия. Рассмотрим, какие процессы протекают в это время в мембране.
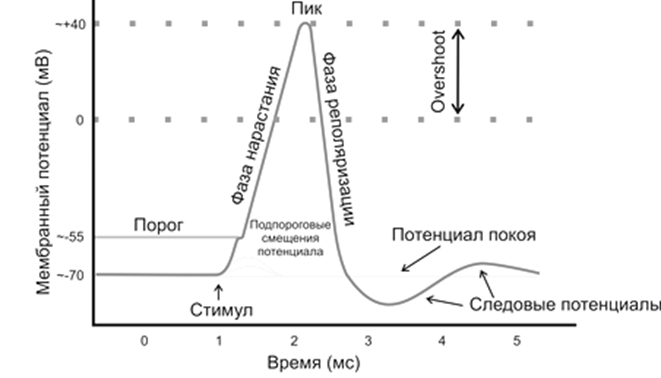 Первая фаза – местная электроотрицательность. В начале развития потенциала действия открываются не все натриевые каналы, ток натрия в клетку невелик. Мембранные потенциал немного уменьшается. Если действовавший раздражитель был слабым (подпороговым), то на этой стадии все и заканчивается. Возбуждение не распространяется за пределы участка, натриевые каналы инактивируются а ионные насосы восстанавливает нарушенное ионное равновесие. Поэтому первую фазу называют местным, или локальным ответом. Если же раздражитель был пороговой или сверхпороговой силы, то местная реакция ткани немедленно переходит во вторую фазу.
Первая фаза – местная электроотрицательность. В начале развития потенциала действия открываются не все натриевые каналы, ток натрия в клетку невелик. Мембранные потенциал немного уменьшается. Если действовавший раздражитель был слабым (подпороговым), то на этой стадии все и заканчивается. Возбуждение не распространяется за пределы участка, натриевые каналы инактивируются а ионные насосы восстанавливает нарушенное ионное равновесие. Поэтому первую фазу называют местным, или локальным ответом. Если же раздражитель был пороговой или сверхпороговой силы, то местная реакция ткани немедленно переходит во вторую фазу.
Вторая фаза – деполяризация мембраны. Если в первой фазе мембранный потенциал снизился на некую определенную величину (10 - 15 мВ), тогда на участке мембраны открываются большинство натриевых каналов. Такая первоначальная величина мембранного потенциала, достаточная для того, чтобы открылись и другие натриевые каналы, называется критическим уровнем деполяризации.
Если единичный канал пропускают примерно 6000 ионов натрия за 1 мс, что создает очень небольшое напряжение тока, то общий натриевый ток равен сумме тысяч очень слабых импульсных токов, обусловленных открытием отдельных натриевых каналов. Входящий ток натрия сначала сдвигает мембранный потенциал до нуля, а затем он возрастакт, но уже с противоположным знаком. Теперь внутри клетки мембрана приобретает положительный заряд (избыток катионов натрия), а наружная поверхность мембраны оказывается электроотрицательной (дефицит натрия). На высоте пика деполяризации величина ПД становится равной 110 - 130 мВ. Это состояние продолжается десятые и сотые доли миллисекунды.
Третья фаза – реполяризация. С этого момента в мембране начинаются восстановительные процессы.
В период реполяризации инактивируются натриевые каналы, что прекращает дальнейшее поступление натрия в клетку. Открываются дополнительные калиевые каналы, благодаря чему больше ионов калия пассивно выходит из клетки. Усиливается работа калий-натриевого насоса, восстанавливающего ионный гомеостаз клетки. Вначале эти процессы протекают быстро, а затем замедляются. Начинаются следовые колебания ПД.
Четвертая фаза ПД – следовая деполяризация. Мембранный потенциал уменьшается по сравнению с 3 фазой, восстанавливается полярность мембраны (знаки зарядов: плюс снаружи, минус – внутри), но он ещё меньше, чем был до возбуждения.
Пятая фаза - следовая гиперполяризация. Как бы по инерции мембранный потенциал продолжает снижаться и оказывается на 5 - 10 мВ меньше первоначального.
Следовые потенциалы могут иметь различную продолжительность в зависимости и от свойств данной ткани, и величины раздражителя, его частоты и длительности состояния возбуждения.
Значение потенциала действия заключается, во-первых, в том, что он является, раздражителем для соседних участков данной клетки, в результате чего возбуждение распространяется по всей ее поверхности, на которые раздражитель не действовал. Во-вторых, ПД становится пусковым механизмом для внутриклеточных процессов: под воздействием электрического тока нерв передает возбуждение, мышца – сокращается, а секреторная клетка выделяет секрет.
Итак, потенциал действия возникает в результате пассивного тока натрия внутрь клетки через натриевые каналы, открывшиеся под влиянием раздражителя. Возбужденный участок клетки оказывается электроотрицательным по отношению к невозбужденному. Пик ПД составляет 110 - 130 мВ. Достигнув пика, мембранный потенциал возвращается к уровню потенциала покоя. Продолжительность одного импульса в разных тканях от 2 до 200 мсек
Дата добавления: 2020-10-25; просмотров: 674;
Поиск по сайту
Узнать еще
- II раздел. Организация работы логопеда в группе для детей с ОНР
- III. СТРУКТУРА И ОРГАНИЗАЦИЯ ДЕЯТЕЛЬНОСТИ
- VI. Организация и финансирование капитального ремонта многоквартирного дома при формировании фонда капитального ремонта на специальном счете
- VII. Организация и финансирование капитального ремонта многоквартирного дома при формировании фонда капитального ремонта у регионального оператора
- Адхократическая организация
- Архитектурный ансамбль: пространственная организация системы площадей, улиц, кварталов, набережных, парков.
- БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
- БЮДЖЕТИРОВАНИЕ В ОРГАНИЗАЦИЯХ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
