Передача возбуждения с нерва на рабочий орган. Синапсы и их свойства. Медиаторы и их роль.
Место контакта аксона с другой клеткой – нервной или мышечной - называется синапсом. Мембрана, покрывающая окончание аксона, называется пресинаптической. Часть мембраны второй клетки, расположенная напротив аксона, называется постсинаптической. Между ними имеется синаптическая щель.
В нервно-мышечных синапсах для передачи возбуждения с аксона на мышечное волокно используются химические вещества –медиаторы (посредники) – ацетилхолин, норадреналин, адреналин и др. В каждом синапсе вырабатывается какой-то один медиатор, и по названию медиатора синапсы называются холинергическими или адренергическими.
В пресинаптической мембране находятся везикулы (пузырьки), в которых накапливаются молекулы медиатора.
На постсинаптической мембране находятся молекулярные комплексы, называемые рецепторами (не путайте с рецепторами – чувствительными нервными окончаниями). В структуру рецептора входят молекулы, «узнающие» молекулу медиатора, и ионный канал. Там же имеется макроэргическое вещество – АТФ, и фермент АТФ-аза, стимулирующий распад АТФ для энергетического обеспечения возбуждения. После выполнения своей функции медиатор должен разрушиться, и в постсинаптическую мембрану встроены гидролитические ферменты: ацетилхолинэстераза, или холинэстераза, разрушающие ацетилхолин и моноаминооксидаза, разрушающая норадреналин.
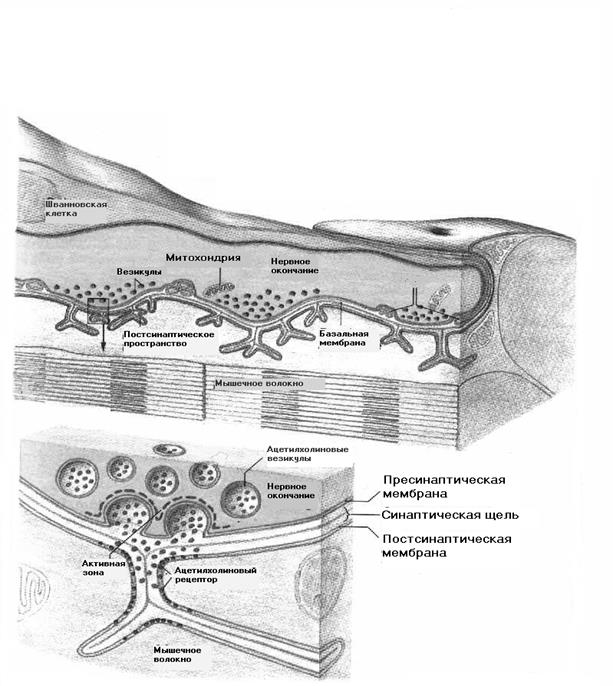 Передача возбуждения в нервно-мышечном синапсе осуществляется следующим образом.
Передача возбуждения в нервно-мышечном синапсе осуществляется следующим образом.
Нервный импульс, достигнув пресинаптической мембраны, вызывает активацию электрозависимых кальциевых каналов и входящий ток ионов кальция в мембрану из синаптической щели. Внутри мембраны кальций способствует выбросу медиатора из везикул в синаптическую щель. Ацетилхолин диффундирует к постсинаптической мембране, и связывается со специальными (рецепторными) участками этой мембраны. В результате происходит активация хемочувствительных натриевых ионных каналов, и входящий натриевый ток вызывает деполяризацию постсинаптической мембраны. Миниатюрные потенциалы на каждом канале суммируются и развивается потенциал действия второй клетки. Его суммарная величина становится раздражителем для соседних участков мышечной клетки.
После окончания действия медиатор разрушается. В результате освобождаются рецепторные комплексы, и ионные каналы снова готовы к восприятию новой порции медиатора. Продукты неполного распада медиатора всасываются в окончание аксона, где происходит ферментативный ресинтез новых молекул медиаторов.
В некоторых холинергических синапсах в состав рецептора входит не натриевые, а калиевые каналы. В результате взаимодействия медиатора с калиевыми каналами увеличивается выход калия из клетки, наступает гиперполяризация мембраны и торможение мышечной клетки. Например, ацетилхолин возбуждает скелетные мышцы, но тормозит гладкие мышцы в пищеварительном тракте и кардиомиоциты. Такими же свойствами обладают и адренергические синапсы. Норадреналин в одних синапсах вызывает возбуждение гладких мышц, а в других – расслабление.
Таким образом, один и тот же медиатор в разных синапсах может вызывать разные эффекты в зависимости от структуры и свойств рецепторного комплекса.
2.5. Механизм мышечного\ сокращения. Энергетическое обеспечение мышечного сокращения
Мышцы являются важнейшими исполнительными органами – эффекторами. По морфологическим и функциональным характеристикам мышцы разделяют на поперечно-полосатые (скелетные) и гладкие (во внутренних органах – сосудах, кишечнике, яйцеводах, мочеточниках и др.). Сердечную мышцу, имеющую ряд структурных и функциональных особенностей, иногда выделяют как третий тип мышц, иногда присоединяют как разновидность к поперечно-полосатым мышцам. Физиологические свойства сердечной мышцы обычно рассматривают при изучении сердечно-сосудистой системы.
Поперечно-полосатые мышцы состоят из отдельных цилиндрических многоядерных клеток или, волокон, которые расположены в общем соединительно-тканном футляре. Диаметр волокон варьирует от 5 до 100 мкм, а длина может достигать у крупных животных более 10 см. Длина гладкомышечных волокон более короткая.
Каждое мышечное волокно состоит из множества параллельно расположенных сократительных субъединиц - миофибрилл, включающих в себя повторяющиеся в продольном направлении блоки – саркомеры, отделенные друг от друга Z- пластинками. В обоих направлениях от Z- пластинки тянутся многочисленные тонкие нити (филаменты), состоящие из белка актина. Между актиновыми нитями располагаются толстые нити, состоящие из белка миозина.
В скелетных мышцах расположение актиновых и миозиновых участков строго упорядочено, поэтому при микроскопии видны чередующиеся темные и светлые участки, откуда и пошло их название - поперечно-полосатые. В гладких мышцах актиновые и миозиновые нити расположены более хаотично, и при рассматривании в микроскоп они не имеют поперечной исчерченности.
 По периметру каждой миофибриллы на уровне Z-пластинки идёт окружённая мембраной поперечная трубочка (Т–трубочка) диаметром около 0,1 мкм. В дополнение к системе Т-трубочек в мышцах есть система, получившая название саркоплазматического ретикулума. Она обволакивает подобно полой манжете отдельно каждую миофибриллу от одной Z-пластинки до другой. Концевые цистерны саркоплазматического ретикулума вступают в контакт с Т-трубочкой. Мембрана мышечного волокна образует регулярные впячивания (трубки диаметром 50 нм), которые находятся вблизи Т-трубочек. Т-трубочки и саркоплазматический ретикулум являются место хранения внутриклеточного кальция. В мембране, покрывающей эту систему, имеются кальциевые каналы и кальциевые насосы. Выход кальция из Т-системы происходит пассивно, через открывающиеся кальциевые каналы, а обратное поступление кальция – из цитоплазмы в Т-систему- осуществляется кальциевым насосом с затратой клеткой энергии АТФ.
По периметру каждой миофибриллы на уровне Z-пластинки идёт окружённая мембраной поперечная трубочка (Т–трубочка) диаметром около 0,1 мкм. В дополнение к системе Т-трубочек в мышцах есть система, получившая название саркоплазматического ретикулума. Она обволакивает подобно полой манжете отдельно каждую миофибриллу от одной Z-пластинки до другой. Концевые цистерны саркоплазматического ретикулума вступают в контакт с Т-трубочкой. Мембрана мышечного волокна образует регулярные впячивания (трубки диаметром 50 нм), которые находятся вблизи Т-трубочек. Т-трубочки и саркоплазматический ретикулум являются место хранения внутриклеточного кальция. В мембране, покрывающей эту систему, имеются кальциевые каналы и кальциевые насосы. Выход кальция из Т-системы происходит пассивно, через открывающиеся кальциевые каналы, а обратное поступление кальция – из цитоплазмы в Т-систему- осуществляется кальциевым насосом с затратой клеткой энергии АТФ.
Помимо актина и миозина, к сократительным белкам мышцы относятся еще два белка – тропонин и тропомиозин. Они расположены между актиновыми и миозиновыми нитями и не позволяют им контактировать, когда мышца находится в состоянии покоя.
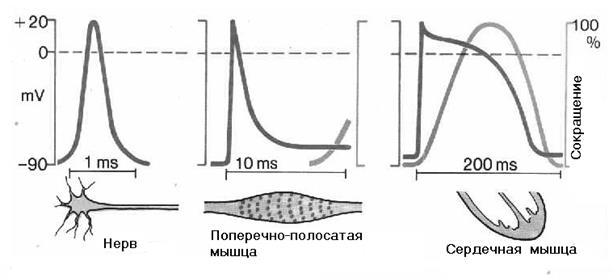
Основными физиологическими свойствами мышц являются возбудимость, проводимость и сократимость.
Возбудимость – возбудимость – это свойство мышечной ткани переходить в активное состояние при действии на нее раздражителя. Адекватными раздражителями для мышц являются потенциалы действия, передающиеся с нервов, а точнее – нервные медиаторы, выделяющиеся в нервных окончаниях. Мышцы могут также возбуждаться при коротком, но достаточном сильном растяжении. Во время возбуждения в мембране мышечного волокна возникает потенциал действия, который служит пусковым механизмом для сокращения.
Проводимость – свойство мембраны мышечного волокна проводить возбуждения от синапса по всей его длине. Передача возбуждения вдоль мембраны осуществляется путем круговых токов – от одного участка к следующему.
В скелетных мышцах возбуждение по каждому мышечному волокну передается изолированно, не переходя на рядом лежащее. Аксон двигательного нервного волокна внутри мышцы разделяется на несколько веточек, каждая из них подходит к отдельному мышечному волокну. Образуется нервно-мышечная моторная единица, моторный (двигательный) нейрон иннервирует несколько мышечных клеток, образуя на каждой из них синапс. Особенности передачи возбуждения в гладких мышцах описано ниже.
Сократимость – свойство мышц сокращаться во время возбуждения. Механизм сокращения состоит в перемещении (протягивания) актиновых нитей вдоль миозиновых к центру саркомера (теория скольжения нитей). Одновременное укорочение тысяч саркомеров в каждой миофибрилле приводит к сокращению всей мышечной клетки.
Сами белковые молекулы при этом не укорачиваются. На толстых миозиновых нитях имеются поперечные мостики, которые при возбуждении клетки изменяют угол наклона и присоединяются к участкам тонких актиновых нитей. При этом контакте активируется фермент АТФ-аза, расщепляющий АТФ с освобождением энергии. Каждый мостик то соединяется и тянет нить, то отсоединяется и ждет «условий» для нового прикрепления.
В покоящейся мышце миозин не может соединиться с актином, поскольку между ними находятся нити белка тропомиозина. Это препятствие устраняется, когда во внутриклеточном пространстве увеличивается концентрация ионов кальция. В присутствии АТФ кальций вызывает конформационные изменения тропомиозина, в результате чего он отклоняется в сторону, и миозин получает возможность контакта с актином. После перемещения актиновой нити на один шаг (20нм) кальций откачивается кальциевыми насосами в саркоплазматический ретикулум, и его концентрация в цитоплазме снижается. Мостик между актином и миозином отсоединяется, а между актиновыми и миозиновыми нитями вновь встраивается тропомиозин. Для продолжения сокращения мышечного волокна требуется новый импульс потенциала действия.
Итак, последовательность процессов при возбуждении и сокращении мышечной клетки следующая.
1. Потенциал действия, возникнув в области синапса, распространяется вдоль мембраны клетки, включая и впячивания мембраны возле Т-трубочек.
2. Потенциал действия с поверхностной мембраны передается на мембрану Т-трубочек и саркоплазматического ретикулума (очевидно, с помощью химического вещества - посредника).
3. Электрический ток, возникший на мембране саркоплазматического ретикулума, активирует Са-каналы, что вызывает выход кальция в миоплазму. Кальций, соединяясь с тропомиозином, вызывает в молекуле этого белка конформационные изменения, в результате чего устраняются препятствия для присоединения миозиновых мостиков к актиновым филаментам, и актиновая нить скользит вдоль миозиновой.
4. Освободившаяся из АТФ энергия затрачивается не только на перемещение (скольжение) нитей актина и миозина, но и на активацию Са-насосов в мембране саркоплазматического ретикулума, вследствие чего кальций переносится в свое депо, концентрация кальция в миоплазме снижается и тропомиозин встает на свое место, разобщая актиновые и миозиновые нити.
Энергетические процессы в мышце. Энергетические процессы в работающей мышце протекают в две фазы – анаэробной и аэробной.
В первую фазу - во время сокращения - происходит распад АТФ до АДФ, остатка фосфорной кислоты и свободной энергии – это основной источник энергии для сокращения мышцы и работы всех ионных насосов (К-Na, Ca). Распад АТФ идет в анаэробных (без участия кислорода) условиях.
Во вторую фазу - во время расслабления мышцы - идет восстановление АТФ: к молекуле АДФ присоединяется остаток фосфорной кислоты и добавляется порция энергии. Источником фосфатной группы, помимо отделившейся от АТФ, является мышечный белок – креатинфосфат. Энергия для аккумулирования в химических связях АТФ освобождается при окислении глюкозы, жирных кислот и других метаболитов углеводного-жирового обмена, что требует участия кислорода (аэробный процесс). Процесс переноса фосфатной группы и энергии, полученной при окислении веществ, на АДФ, и ресинтез АТФ называется окислительным фосфорилированием.
Таким образом, в работающей мышце с очень большой скоростью происходит смена аэробных и анаэробных процессов. Запас АТФ при этом не уменьшается (АТФ – АДФ – АТФ), а затрачиваются внутриклеточные запасы гликогена как источника глюкозы, и другие вещества, как образующиеся в клетке в процессе окисления, так и поступающие с током крови.
При длительной интенсивной работе мышце не хватает кислорода для окисления продуктов неполного распада окисляемых веществ, в ней накапливаются такие вещества, как молочная, пировиноградная, уксусная, фосфорная кислоты, креатинин. Это вызывает нарушение внутриклеточного гомеостаза, снижение рН и работоспособность мышцы уменьшается вплоть до полного отказа сокращаться, что называется утомлением.
Однако не вся освобождающаяся при работе мышцы энергия затрачивается на обеспечение ее сокращения. Бóльшая часть энергии (до 80%) переходит в тепловую, она идет на поддержание постоянной температуры тела, а ее излишек рассеивается в окружающей среде. Коэффициент полезного действия мышц довольно высок, он составляет 20-30%.
Сократительные свойства мышц. Сокращение мышцы может происходить в двух режимах: изотоническом и изометрическом (изо – равный, одинаковый, тонус – напряжение, метр – длина).
При изотоническом сокращении мышца укорачивается, но ее напряжение, почти не изменяется. Например, такое сокращение бывает при небольшой величине поднимаемого груза, или при сокращении мышцы без груза. При изометрическом режиме длина мышцы не меняется, а развивается внутреннее напряжение. Примером такого сокращения может быть большое напряжение мышцы при поднятии чрезвычайно большого, непосильного груза.
В зависимости от частоты раздражения различают два вида сокращений мышц: одиночное и тетаническое. Одиночное сокращение возникает при однократном возбуждении мышечных волокон. Во время одиночного сокращения прикрепление миозиновых мостиков к актиновым филаментам быстро устраняется вследствие того, что ионы кальция возвращаются в саркоплазматический ретикулум. Поэтому напряжение, на которое способна сократительная система, в одиночном мышечном сокращении полностью не реализуется.
При более частых раздражениях мышца не успевает расслабиться и наблюдается длительное сокращение без расслабления. Такие сокращения мышцы называются тетаническими, или тетанусом. При тетанических сокращениях сила сокращения мышцы увеличивается по сравнению с одиночным сокращением. Это обусловлено тем, что ионы кальция после каждого импульса не успевают выкачиваться из цитоплазмы, и миофиламенты (актиновые и миозиновые) продолжают скольжение, усиливая сокращения.
В зависимости от частоты потенциалов действия возникают разные степени слияния (суммации, сложения) одиночных сокращений, и развиваются разные формы тетанического напряжения. При более редких раздражениях возникает зубчатый тетанус, при более частых – гладкий. В первом случае заметны «вздрагивания» мышцы при каждом пике сокращения, во втором - они сливаются. Примерами зубчатого тетануса может быть терморегуляционная дрожь при холоде, а гладкого тетануса – сокращения мышц при длительном удержании груза.
Сокращение мышцы сопровождается ее работой. Принято различать статическую работу - удержание позы (определенного положения тела в пространстве), противодействие силе притяжения Земли, и динамическую - перенос груза, передвижение тела животного.
Динамическую работу измеряют произведением массы перемещаемого груза на пройденное расстояние. При маленьком грузе выполняемая работа невелика, но и при очень большом грузе, когда мышцы не могут поднять его на большую высоту, работа тоже оказывается небольшой. Максимальную работу мышцы совершают при средней величине груза (правило средней нагрузки). Эту закономерность следует учитывать и при физической работе, и при тренировках.
Сила мышцы характеризуется максимальным грузом, который мышца может поднять хотя бы на небольшую высоту. Она зависит от количества мышечных волокон в мышце, от количества и толщины миофибрилл. Высота поднятия груза обусловлена длиной мышечных волокон. Поэтому перистые мышцы, в которых мышечные волокна расположены под разными углами, и их много, более сильные, чем мышцы с параллельными волокнами, но они сокращаются с небольшой амплитудой. Таковы, например, икроножные мышцы. Зато мышцы с параллельными волокнами, не обладая большой силой, могут сокращаться на бóльшую длину и выполнять, например, мáховые движения (портняжная мышца).
Растяжимость, эластичность и пластичность мышц – тоже являются их сократительными свойствами. Растяжимость – способность мышц пассивно растягиваться, удлиняться под воздействием груза. Эластичность – свойство мышц возвращаться к первоначальной длине после снятия растяжения. Пластичность – свойство мышц сохранять приданную форму после растяжения. Скелетные мышцы характеризуются высокой эластичностью, а гладкие мышцы, особенно мышцы матки, мочевого пузыря, желудка – очень пластичны, и могут долгое время оставаться в растянутом состоянии после опорожнения.
Физиологические особенности гладких мышц обусловлены особенностями их строения. Они не содержат упорядоченных слоёв актиновых и миозиновых филаментов, поэтому при световой микроскопии не имеют поперечной исчерченности, отсюда их название – гладкие.
Гладкие мышцы иннервируются вегетативными нервами, симпатическими и парасимпатическими. Нервные окончания не образуют синапсов. Медиаторы - ацетилхолин, норадреналин - выделяются в расширенных участках нервного волокна, и они оказывают влияние одновременно на несколько мышечных клеток. Гладкомышечные клетки объединены друг с другом через плотные контакты, которые обеспечивают, подобно электрическим синапсам, распространение электрического тока от клетки к клетке, поэтому в гладких мышцах возбуждение может передаваться от одного мышечного волокна на соседнее.
Гладкие мышцы менее возбудимы, сокращаются и расслабляются значительно медленнее, чем поперечно-полосатые. Они имеют более продолжительные и латентный период, и период сокращения. Поэтому гладкие мышцы могут длительное время находиться в состоянии возбуждения под влиянием редких импульсов. Например, мышцы сосудов, сфинктеров находятся в повышенном тонусе в течение почти всей жизни организма, не утомляясь, с небольшой затратой энергии.
В процессе возбуждения поверхностной мембраны имеет значение не только входящий натриевый ток, но и кальциевый, осуществляющийся через кальциевые каналы. С помощью Ca-насоса ионы кальция постоянно выводятся через мембрану, в результате чего внутриклеточная концентрация этого иона в покоящейся мышце сохраняется на достаточно низком уровне.
Гладкие мышцы могут легко растягиваться под воздействием какой-либо силы (заполнением полых органов), они очень пластичны. Поэтому заполненный желудок или переполненный мочевой пузырь могут длительное время находиться в растянутом состоянии, не сокращаясь. Эластичность гладких мышц не совершенна, после опорожнения органа они не сразу возвращаются в начальное состояние.
В некоторых органах (кишечник, мочеточник) гладкомышечные клетки обладают автоматией, то-есть способностью к спонтанной деполяризации мембраны. В этих органах наблюдаются более или менее ритмичные сокращения без участия нервной системы или гуморальных (химических) воздействий. Например, участок кишечника, изолированный от организма и помещенный в физиологический раствор, продолжает перистальтические сокращения.
Дата добавления: 2020-10-25; просмотров: 781;
Поиск по сайту
Узнать еще
- I. Товар и его свойства.
- Автоматизация и контроль. Диспетчеризация
- Автоматическими регуляторами возбуждения (АРВ)
- Адренэргнические синапсы
- АЛГОРИТМ И ЕГО СВОЙСТВА. Определение алгоритма
- Аллергические реакции 1 типа (реагиновые). Стадии, медиаторы аллергии 1 типа, механизмы их действия. Клинические проявления (анафилактический шок, атопические реакции).
- Аритмии вследствие нарушения возбудимости и проведения возбуждения в сердечной мышце.
- Асинхронная передача данных

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
